Флористическое деление суши и океана
Флористическое деление суши
Каждая страна, каждая область суши имеет свою, характерную для нее флору, т. е. свой набор семейств, родов и видов, который более или менее отличается от флоры других стран. Флоры (как и фауны) никогда не бывают совершенно одинаковыми в двух странах или в двух разных областях одной и той же страны.
Эти различия отчасти объясняются разнообразием геологических, орографических, почвенных и особенно климатических условий. Так, бананы, ананас или тропические пальмы не могут произрастать в областях с холодным или даже умеренным климатом, а северная береза не встречается в тропических странах.
Но в то же время в тропических областях разных стран и особенно разных материков наблюдаются большие различия в составе флоры, причем даже в тех случаях, когда почвенные и климатические условия очень сходны. Так, семейство бромелиевых (Bromeliaceae), куда входит ананас, встречается почти исключительно в тропической и отчасти субтропической Америке (только один вид в тропической Западной Африке), в то время как другое тропическое семейство — пандановых (Pandanaсеае) совершенно отсутствует в Америке и приурочено к тропическим и отчасти внетропическим областям Старого Света. Таких примеров можно привести множество.
Эти факты говорят о том, что характер флоры данной области нельзя объяснить только почвенными и климатическими условиями. Очевидно, флористические различия объясняются и другими факторами. Таковы географическая изоляция, миграция и дифференциальное вымирание (полное или частичное) отдельных флористических комплексов или целых флор.
В результате географической изоляции многие области Земли характеризуются очень своеобразной флорой, в составе которой много эндемичных (произрастающих только на данной территории) видов, родов и даже семейств.
Исключительное своеобразие и высокий эндемизм флоры Австралии, Мадагаскара и Новой Каледонии объясняются очень длительной их изоляцией. Географическая изоляция характерна не только для материков и островов, но и для отдельных высоких гор или целых горных систем (например, Альпы, Кавказ или Гималаи), пустынь, окруженных более влажными территориями, и вообще для любой области, окруженной теми или иными барьерами, служащими преградой для распространения растений.
Одним из важнейших факторов, изменяющих состав флоры любой страны, является расселение, или миграция, растений, т. е. их распространение из одной области в другую. Процесс миграции происходит с разной скоростью, то очень быстро, то, наоборот, очень медленно и постепенно.
В некоторых случаях из одной области в другую могут переноситься целые растения. Это относится к одноклеточным и небольшим водным растениям (водоросли, ряски и др.) или к растениям типа перекатиполе, например Ceratocarpus arenarius из семейства маревых (Chenopodiaceae). Обычно переносится не все растение, а лишь определенная его часть: споры, семена и плоды, соплодия, сеянцы (у мангровых растений), выводковые почки (у печеночника маршанции), клубеньки и пр.
Для обозначения частей растения, служащих для распространения, шведский ботаник Сернандкр (1927) предложил специальный термин — диаспора (от греч. diaspeiro — разбрасываю вокруг, рассеиваю). Синонимами диаспоры являются единицы распространения: зачаток, пропагула, гермула, мигрула и диссемула.
Морские течения, ветер и животные (особенно птицы) могут способствовать быстрому распространению диаспор иногда на очень далекие расстояния. Изолированные вулканические острова, возникающие в океане далеко от материков, могут заселяться только таким путем. Именно так заселялись, например, Гавайские острова. Широкое распространение многих пресноводных растений обязано главным образом птицам, а распространению мангровых растений способствуют морские течения.
В большинстве случаев, однако, диаспоры распространяются более медленно и постепенно, и, как правило, расселение идет по определенным путям и направлениям, которые называются миграционными путями.
Миграционный путь редко представляет собой вполне удобную трассу для беспрепятственного распространения растений. Обычно на нем встречаются различного рода преграды, затрудняющие расселение. Наиболее удобным миграционным путем является путь, названный известным палеонтологом и биогеографом Дж. Симпсоном (1965) «коридором». «Коридор» представляет собой миграционную трассу, вдоль которой возможно массовое распространение многих видов или даже целых флор из одной области в другую. Типичным «коридором» служило существовавшее в третичное время море Тетис.
«Коридорами» являются также речные долины или непрерывные горные цепи, как, например, Гималаи.
В отличие от «коридора» Симпсон называет «фильтром» миграционный путь, по которому расселение происходит очень избирательно. Для одних видов растений «фильтр» представляет собой вполне удобный миграционный путь, в то время как для многих других он является непреодолимым барьером. В результате не все элементы флоры данной области могут мигрировать в другую область. Для лесных растений «фильтром» может быть степь, а для степных — пустыня. Как отмечает Симпсон. межконтинентальные мосты суши отличаются сильно выраженным «фильтровым» характером. Таким «фильтровым мостом» является, например, мост, соединяющий Северную Америку с Южной. Другим, более близким нам, примером является кавказский перешеек, служащий «фильтровым мостом» между южнорусскими и переднеазиатскими флорами.
Основным, и притом широко распространенным, типом миграционного пути является «ступенчатый» путь.
Он представляет собой прерывистый путь, состоящий из отдельных «ступенек», подобно каменной переправе через речку. Это или цепь островов (архипелаг типа Алеутских островов), или цепь изолированных гор, отделенных глубокими долинами, или цепь озер. В лесостепной зоне «ступенчатым» миграционным путем является ряд лесных участков, отделенных степными пространствами. В отличие от «коридоров» «ступенчатый» путь характеризуется значительно меньшей скоростью миграции и большей избирательностью (в этом отношении он приближается к «фильтру»). Наличие многих островных «ступенек» особенно характерно для Тихого океана, что играло очень большую роль в истории флоры Тихоокеанского бассейна.
В результате совместного действия трех основных факторов эволюции, миграции и вымирания (частичного или полного) - возникла чрезвычайно сложная и мозаичная картина современной флоры Земли. Эволюция не происходит без миграции, хотя бы очень ограниченной, а миграция, когда она происходит медленно и постепенно, обычно сопровождается эволюционными изменениями, хотя бы на подвидовом уровне. Вымирание обычно также представляет собой процесс медленный и постепенный. Но во многих случаях благодаря резким изменениям климатических и других физико-географических условий, а иногда и по чисто биологическим причинам происходит резкое изменение или даже катастрофическое уничтожение целых флор. Так, например, из-за оледенений, происходивших в четвертичном периоде, на значительных пространствах Северной Евразии была уничтожена богатая лесная третичная растительность, отдельные участки которой сохранились лишь в некоторых убежищах (рефугиумах) третичной флоры, например в Колхиде и Талыше. После уничтожения прежней флоры всегда рано или поздно следует интенсивная иммиграция, и тогда образуется новая флора, иногда смешанная, возникшая из разных источников. Ярким примером может служить остров Кракатау близ Явы, на котором вулканическое извержение в 1883 г. полностью уничтожило весь растительный покров, но уже в 1933 г. на нем успешно произрастали растения 271 вида.
Флора каждой страны слагается из: уцелевших и неизменившихся или слабо изменившихся древних форм, обитавших здесь еще до четвертичного периода; видов, представлявших непосредственный результат изменения третичных форм под влиянием условий жизни в данной стране; видов, переселившихся в позднейшую эпоху.
В своей классической работе «Опыт истории развития флоры южной части Восточного Тянь-Шаня» (1888) А. Н. Краснов обозначает флору всякой данной страны формулой
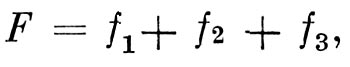
где F совокупность всех ныне живущих видов; f1, f2, f3 — три указанных выше элемента. На основании относительного преобладания того или иного из этих трех элементов А. Н. Краснов различает три типа флор.
Первый тип флоры, который известен под названием реликтового, характерен для Канарских островов, Японии, Китая, некоторых частей Северной Америки и многих других стран. Благоприятные орографические и климатические условия способствовали сохранению большого процента мало изменившихся древних, реликтовых форм, уже давно произрастающих в данной области. Флоры подобного типа наиболее приближаются к идеальной формуле F = f1.
«Большая часть Западной Европы, — пишет А. Н. Краснов, — может быть представительницей другого типа. Ход исторических событий был здесь особенно бурный, непостоянный. Подобно тому как древняя греко-римская цивилизация погибла под напором вторгнувшихся варваров — и от нее до нас дошли лишь жалкие остатки, так же точно, за исключением немногих представителей, под влиянием холода ледникового периода погибли роскошные третичные формы — и новая, арктического характера флора заняла их место. Но господство ее продолжалось лишь некоторое время; вскоре, вследствие обусловленного различными признаками смягчения климата, древняя флора стала теснить арктическую и во многих местах вытеснила ее почти совершенно. Таким образом, Европа была страной постоянного движения форм, страной переселений и, в то время как в Японии и на Мадейре за весь длинный период времени, протекший от конца третичного периода до настоящей минуты, почва была одета флорой приблизительно сходного характера, в Европе можно найти много пунктов, флора которых за этот период изменилась радикально.
Таким образом, тут флора менялась дважды. И хотя ныне существующая и сходна с древнетретичной, но она все же стоит от нее так же далеко, как далеки итальянцы от своих славных предков — римлян. Флора здесь стремится приблизиться к идеальному случаю F = f3». Флоры подобного типа были названы миграционными.
Флора Средней Азии может служить, по мнению А. Н. Краснова, типом третьего рода флор. «С давно прошедших времен и до настоящей минуты климат страны постоянно изменяется, но изменения эти происходят все в одну и ту же сторону. Как в странах первого типа, так и здесь ледниковый период и другие климатические изменения хотя и давали себя чувствовать более или менее сильно, но не могли, однако, изменить общего хода событий — в данном случае, например, постепенного усыхания, усиления континентальности. Постепенное и все усиливающееся действие одних и тех же факторов на флору не могло не вызвать в ней соответствующих приспособлений и изменений.
В то время как в Западной Европе, при постоянных изменениях климата и переселениях, как полагают Христ и Де Кандоль, не образовалось со времен третичного периода ни одного хорошего вида, здесь, под влиянием векового и постоянно усиливающегося действия одних и тех же факторов на миллионы поколений, можно ожидать — на основании многих данных — противного. Здесь, где флора ближе всего к идеальному F = f2, легче, чем где-либо, путем сравнения ее с третичной или флорами типа F = f1 изучать историю перерождения палеоарктической флоры в современную. Хотя и здесь формула флоры будет F = f1 + f2 + f3 но при преобладании /2 остальные, менее многочисленные элементы легче могут быть выделены». Такие флоры можно назвать трансформационными.
Эти три установленных А. Н. Красновым типа флор редко встречаются в чистом виде. и в природе мы обычно имеем дело с различными их комбинациями.
Сравнительное изучение флор разных стран привело к необходимости создать флористическую систему, т. е. разделить земной шар на естественные флористические единицы. Эти попытки предпринимались уже давно. Одна из первых, и притом для своего времени чрезвычайно удачная, принадлежит датскому ботанику Скоу (1823). Скоу разделил флору земного шара на 25 царств, некоторые из которых, в свою очередь, подразделяются на провинции (термины, до сих пор принятые в ботанической географии). В таком делении Скоу, как и последующие авторы, учитывал степень эндемизма, т. е. число таксонов, произрастающих только на данной территории. Он считал, что для выделения той или иной территории в особое флористическое царство необходимо, чтобы по крайней мере половина видов и четверть родов были эндемичны. Кроме того, флористическое царство должно характеризоваться также эндемичными семействами или хотя бы семействами, характеризующимися разнообразием в пределах территории царства. Современный подход к флористическому районированию во многом близок к принципам, сформулированным много лет назад Скоу.
Впоследствии разработкой флористической системы земного шара занимался ряд крупных ботаников, из которых наиболее крупный вклад сделал А. Энглер.
Все последующие системы, в том числе флористическая система автора этих строк, представляют собой дальнейшее развитие флористического деления А. Энглера, первый вариант которого был опубликован в 1882 г., а последний — в 1924 г.
Существует некоторое, хотя и не очень строгое, соответствие между рангом единиц флористической системы (в современной ботанической литературе единицы флористической системы называются фитохорионами): царствами, областями, провинциями и округами — и таксономическим рангом тех систематических единиц (таксонов), по которым устанавливается различие между флористическими территориями. Так, флористические царства характеризуются определенным, в некоторых случаях довольно значительным, семейственным эндемизмом и очень высоким родовым эндемизмом, в то время как флористические области устанавливаются о'бычно на основании наличия эндемичных родов и достаточно высокого видового эндемизма. Флористические области обычно представляют собой крупные очаги видообразования и часто характеризуются эндемичными секциями и подсекциями даже в крупных неэндемичных родах. Флористические провинции далеко не всегда характеризуются эндемичными родами,- и их видовой эндемизм значительно менее высокий, чем у областей. Наконец, округи обладают главным образом подвидовым и лишь слабо выраженным видовым эндемизмом. Округи — это самые молодые фитохорионы, происхождение которых относится к геологически недавнему времени, очень часто к наиболее поздним эпохам четвертичного периода.
Флора суши земного шара подразделяется на 6 флористических царств (цветная карта 2 на вклейке).
I. Голарктическое царство
Голарктическое царство — самое крупное из всех и занимает больше половины всей суши. Оно охватывает всю Европу, северную внетро-пическую Африку, всю внетропическую Азию и почти всю Северную Америку. Несмотря на огромную протяженность его территории и ее разъединенность на европейскую и североамериканскую части, флоры отдельных областей царства тесно связаны между собой и имеют общее происхождение.
В составе голарктической флоры около 40 эндемичных семейств, из которых назовем Calycanthaceae, Glaucidiaceae, Hydrastidaceae, Circaeasteraceae, Hypecoaceae, Trochodendraceae, Tetracentraceae, Cercidiphyllaceae, Eupteleaceae, Platanaceae (один вид в Индокитае), Eucommiaceae, Leitneriaceae, Paeoniaceae, Stachyuraceae, Davidiaceae, Helwingiaceae, Adoxaceae, Scheuchzeriaceae. Широко представлены семейства магнолиевых (Magnoliaceae), лавровых (Lauraceae), лютиковых (Ranunculaceae), барбарисовых (Berberidaceae), гамамелидовых (Hamamelidaceae), буковых (Fagaceae), березовых (Betulaceae), ореховых (Juglandaceae), гвоздичных (Caryophyllaceae), маревых (Chenopodiaceae), гречишных (Polygonaceae), свинчатковых (Plumbaginaceae), чайных (Theaceae), ивовых (Salicaceae), крестоцветных (Brassicaceae), вересковых (Ericaceae), первоцветных (Primulaceae), мальвовых (Malvaceae), молочайных (Euphorbiaceae), волчниковых (Thymelaeaceae), розоцветных (Rosaceae), камнеломковых (Sa-xifragaceae), бобовых (Fabaceae), кизиловых (Cornaceae), аралиевых (Araliaceae), зонтичных (Apiaceae), крушинных (Rhamnaceae), горечавковых (Gentianaceae), бурачниковых (Boragi-naceae), норичниковых (Scrophulariaceae), губоцветных (Lamiaceae), колокольчиковых (Cam-panulaceae), сложноцветных (Asteraceae), лилейных (Liliaceae), ирисовых (Iridaceae), орхидных (Orchidaceae), ситниковых (Juncaceae), осоковых (Сурегасеае) и злаков (Роасеае). Из хвойных широко представлены сосновые (Pinaceae) и кипарисовые (Cupressaceae). Папоротники представлены главным образом семействами аспидиевых (Aspidiaceae) и многоножковых (Polypodiaceae). В большинство этих семейств входит много эндемичных голарктических родов и множество эндемичных видов.
Голарктическое царство подразделяется на три подцарства — Бореальное, Древнесреди-земноморское и Мадреанское.
А. Бореальное подцарство
Бореальное подцарство — самое обширное из трех подцарств голарктической флоры. В то же время оно характеризуется наиболее богатой флорой, в составе которой гораздо больше эндемичных семейств и родов, чем в двух других подцарствах. Для некоторых областей Бореаль-ного подцарства характерно значительное число древних и примитивных семейств и родов.
1. Циркумбореальная, или Евросибирско-Канадская, область.
Это самая крупная флористическая область суши, значительная часть которой находится на территории Советского Союза. Сюда входят Европа (за исключением частей, относящихся к Средиземноморской области), Северная Анатолия, Кавказ (за исключением аридных частей и Талыша), Урал, Сибирь, Дальний Восток (за исключением юго-восточной части по среднему течению Амура), Камчатка, Северный Сахалин, северные Курилы, Алеутские о-ва, Аляска, большая часть Канады и небольшая часть США (в районе оз. Верхнего и в горах к северо-востоку от оз. Онтарио). В большинстве флористических систем евразийская и Северо-Американская части области относятся к разным областям и лишь немногими, как, например, Браун-Бланке (1923), признается единая Евросибирско-Североамериканская область. Для этой области наиболее характерны широколиственные и хвойные леса, а также тундра и степи.
Из хвойных наиболее распространены виды: сосны (Pinus), ели (Picea), пихты (Abies) и лиственницы (Larix), а в Канаде также цуга (Tsuga) и туя (Thuja). Из многочисленных лиственных пород наиболее характерны для этой области виды: дуба (Quercus), бука (Fagus), березы (Betula), ольхи (Alnus), клена (Acer), граба (Carpinus), тополя (Populus), ивы (Salix), ясеня (Fraxinus), вяза (Ulmus), липы (Tilia), грецкого ореха (Juglans), каркаса (Celtis), острии (Ostrya), кизила (Cornus). Из других древесных растений характерны также виды: сливы и вишни (Prunus), ежевики (Rubus), боярышника (Crataegus), груши (Pyrus), яблони (Mains), рябины (Sorbus), спиреи (Spiraea), клекачки (Staphylea), рододендрона (Rhododendron), жимолости (Lonicera), калины (Viburnum), бузины (Sambucus), крушины (Rhamnus), брусники и черники (Vaccinium). Наряду с лесами очень характерны луга, которые в южных районах Европейской части СССР и Сибири переходят в степь. Для северных районов характерны обширные болота и тундра. В горах лесная растительность образует пояса, верхний из которых обычно состоит из хвойных лесов. Выше лесного пояса расположен пояс высокогорной (субальпийской и альпийской) растительности, очень богатой эндемичными формами.
Во флоре Циркумбореальной области нет эндемичных семейств, но здесь много эндемичных или почти эндемичных родов, в том числе крестоцветные Lunaria (Средняя и Юго-Восточная Европа), Borodinia (Восточная Сибирь), Gorodkovia (Северо-Восточная Сибирь), Redowskia (Якутия), род Soldanella из семейства первоцветных (Южная и Средняя Европа), зонтичные Physospermum, Astrantia и Thorella (последний род только в Европе), род Pulmo-naria (медуница) из семейства бурачниковых, род Erinus из семейства норичниковых (Пиренеи, Апеннины и Альпы), три рода из преимущественно тропического семейства гесне-риевых — Ramonda (Центральные и Восточные Пиренеи и прилегающие горы Северо-Восточной Испании, Балканский п-ов) и Haberlea и Jankaea (оба на Балканском п-ове), род Stratiotes (телорез) из семейства водокрасовых, сложноцветное Telekia (от Средней Европы до Кавказа), а также много эндемичных видов. Особенно много эндемичных видов в горных районах -в Пиренеях, Альпах, Карпатах, на Кавказе, в горах Сибири и Канады.
Обширная Циркумбореальная флористическая область расчленяется на ряд провинций, объединенных особенностями состава флоры и своим эндемизмом. Учитывая, что большая часть Циркумбореальной области лежит в пределах территории СССР, мы считаем нелишним несколько подробнее охарактеризовать ее и дать перечень всех провинций, в том числе лежащих за пределами СССР.
Арктическая провинция. В эту циркумполярную провинцию входят Гренландия, Исландия, Фарерские о-ва, Ян-Майен, северные безлесные районы Норвегии, Финляндии, Европейской части СССР и Сибири (включая все Чукотское нагорье и низовья Анадыря), Медвежий, Шпицберген, Земля Франца-Иосифа, все острова сибирских морей, безлесные районы Северной Америки, все острова к северу от Североамериканского материка; в Беринговом море Арктическая провинция доходит на юг до о-вов Прибылова.
Флора этой провинции очень бедна и насчитывает немногим более 1000 видов. При этом во флоре Гренландии около 400 видов, Исландии — 375 видов, востока Болыпеземельской тундры — 468 видов, Новой Земли — 200 видов, Земли Франца-Иосифа — только 37 видов. Флористически относительно наиболее богата Чукотка, насчитывающая около 800 видов. Это-объясняется тем, что в течение плейстоцена она не подвергалась сплошному оледенению. Эндемичных родов в Арктической провинции почти нет, если не считать маленького рода Dupontia (всего 2 вида) из семейства злаков. Но эндемичных видов и подвидов в арктической флоре более 100, главным образом на Чукотке и Аляске. Среди них имеются лютик (Ranunculus sabinei), мак (Papaver polare), несколько видов ивы (Salix arctica и др.), виды крестоцветного рода Вгауа, виды мытника (Pedicularis), злаки (Colpodium vahlianum, С. wrightii, Puccinellia Angustatа) и др.
Арктическая флора сравнительно очень молода. Хотя первые этапы ее развития относятся, вероятно, к началу четвертичного (антропогенового) периода, но полностью она сформировалась лишь в течение послеледникового, или голоценового, времени, т. е. за последние 10 тыс. лет. При этом очень большую роль сыграла иммиграция видов, произраставших ранее вокруг пояса сплошного оледенения.
Растительный покров Арктической провинции беден и почти лишен деревьев. На юге, особенно в более многоснежных районах, характерны кустарниковые сообщества. Кустарники представлены главным образом арктическими видами ивы и березы. Очень характерны кустарнички из семейства вересковых (виды Vaccinium и Arctostaphylos) и травянистые растения из семейства осоковых — виды пушицы (Eriophorum) и осоки (Сагех). В более северных широтах растительность состоит из практически одноярусных сообществ. Особенно характерны виды куропаточкой травы (Dryas) из семейства розоцветных и некоторые стелющиеся ивы (Salix polaris и др.), а также многолетние травянистые растения из семейства злаков, крестоцветных и гвоздичных. Очень характерны виды камнеломки (Saxifraga) из семейства камнеломковых, мытника (Pedicularis) из семейства норичниковых, лапчатки (Potentilla) из семейства розоцветных и оксирия (Oxyria digyna) из семейства гречишных. Однолетних трав очень мало, а нередко они полностью отсутствуют.
Атлантическо-Европейская провинция. Простирается от северо-западных и северных частей Пиренейского п-ова до Юго-Западной Норвегии и включает большую часть Франции (Армориканский массив, Аквитанский и Парижский бассейны, Центральный массив), низовья Рейна, большую часть Северо-Германской низменности, Ютландию, всю Великобританию и Ирландию с прилегающими островами. Ее восточная граница приблизительно соответствует восточной границе падуба (Ilex aquifolium) в Западной Европе. В Северо-Восточной Европе ряд атлантических видов достигает Финского залива. Атлантические виды распространены в Парижском бассейне до Парижа, иногда даже до Шампани и захватывают низменные части Центрального массива.
Эндемичные роды Petrocoptis из семейства гвоздичных (Пиренеи) и Thorella из семейства зонтичных (Португалия и Франция); имеется несколько десятков эндемичных видов, из которых отметим хохлатку (Corydalis claviculata), европейский утесник (Ulex europaeus) и ряд близких к нему видов, английский дрок (Genista anglica), щетинковидный луговик (Deschampsia setacea). Характерно наличие целого ряда теплолюбивых растений, что объясняется мягкостью зимы.
Леса образованы главным образом листопадными деревьями, дубами и буком, из которых первые преобладают на юге, а вторые — в центральной части, на севере и востоке. В Западной и Центральной Франции (Нормандия) появляется также белая пихта (Abies alba). Нередок каштан (Castanea sativa). Характерно наличие в подлеске вечнозеленых растений: падуба (Ilex aquifolium), иглицы (Ruscus aculeatus). Обычен плющ (Hedera helix).
Наряду с лесами распространены кустарниковые заросли и верещатниковые пустоши, для которых характерны вересковые (Calluna и Erica) и безлистные бобовые (Ulex europaeus, Genista anglica и Cytisus scoparius).
Централъноевропейская провинция простирается от Центрального массива, Лотарингского плоскогорья, массива Арденны, Рейнских Сланцевых гор, Юго-Восточной Норвегии, Южной Швеции и Аландских о-вов до побережья Финского залива (включая побережье Карельского перешейка и окрестности Ленинграда). Восточная граница проходит через Латвию, Юго-Восточную Литву (к югу от окрестностей Вильнюса) и, огибая с востока Беловежскую пущу, идет западнее Львова к югу на Днестр и далее через окрестности Черновцов и далее по Пруту спускается к Нижнедунайской низменности.
Типичная среднеевропейская флора обычно не идет на восток дальше Беловежской пущи и восточных предгорий Карпат, но отдельные элементы доходят до Валдайской возвышенности и даже до Свердловска, причем выклинивание этих элементов происходит довольно постепенно, что сильно затрудняет определение восточной границы провинции. В общем восточная граница более или менее соответствует общей восточной границе распространения таких растений, как тис (Tuxus baccata, одно из наиболее крупных в Европе насаждений около Коломыя в Прикарпатье), белая пихта (доходит до Беловежской пущи), бук (Fagus sylvatica), дуб (Quercus petraea), плющ (Hedera helix), звездовка (Astrantia major), европейский равноплодник (Isopyrum thalictroides), вербейник (Lysimachia nemorum), герань (Geranium phaeum), шалфей (Salvia glutino-sa), скополия (Scopolia carniolica), лук (Allium vineale), ячменоволоснец (Hordelymus europaeus), осока (Сагех distans) и др.
Эндемизм флоры этой провинции составляет 10—15%, а в горных районах — до 20%, причем имеется ряд монотипных эндемичных родов, как крестоцветное Rhizobotrya (Южные Альпы), роды Hacquetia и Hladnikia из зонтичных, сложноцветное Berardia (только Альпы) и др.
Из центральноевропейских эндемичных видов, иногда проникающих в соседние провинции, отметим борец (Aconitum paniculatum), гвоздику (Dianthus alpinus. Альпы), рододендрон (Rhododendron hirsutum, Альпы), солда-неллу (Soldanella carpatica, Западные Карпаты), шиповник (Rosa abietina), камнеломку (Saxifraga muscoides, Альпы), клевер (Trifolium saxatile), бутень (Chaerophyllum villarsii), борщевик (Heracleum carpaticum, Восточные Карпаты), сирень (Syringa josikaea, горы Тран-сильвании и Украинские Карпаты), валериану (Valeriana tripteris), колокольчики (Campanula zoysii, Альпы; С. carpatica, Карпаты), медуницу (Pulmonaria filarzkyana, Карпаты), эдельвейс (Leontopodium alpinum), нарциссы (Narcissus poeticus, Альпы; N. angustifolius, Альпы и Карпаты), кукушник (Gymnadenia albida), осоку (Carex carvula), вейник (Calamagrostis villosa).
Характерны хвойные: белая пихта, сосны (Pinus sylvestris, P. mugo и Р. cembra), ель (Picea abies) и тис (Taxus baccata).
Из лиственных древесных пород характерны бук, дубы (Quercus petraea, Q. robur и Q. pubescens), ольха (Alnus viridis и A. glutino-sa), березы (Betula pendula и В. pubescens), граб (Garpinus betulus), лещина (Corylus avellana), клены (Acer campestre, A. platanoides и A. pseudoplatanus), ясень (Fraxinus excelsior), вязы (Ulmus glabra, U. minor и U. laevis), липы (Tilia platyphyllos и Т. cordata), крушина (Rhamnus catharticus), боярышник (Crataegus), слива (Prunus), груша (Pyrus), кизил (Cornus mas и Svida sanguinea), бузина (Sambucus racemosa) и жимолость (Lonicera nigra).
Центральноевропейская провинция подразделяется на несколько подпровинций — Податлантическую, Апеннинскую, Альпийскую, Карпатскую и др., которые многими авторами, в том числе Энглером и Дильсом, рассматриваются как самостоятельные провинции.
Иллирийская (Балканская) провинция охватывает Балканский п-ов, за исключением тех его частей, которые относятся к Централь-ноевропейской и Эвксинской провинциям и к Средиземноморской области.
Эта провинция обладает довольно высоким эндемизмом, причем здесь имеются даже два эндемичных рода из семейства геснериевых — Haberlea (Центральная Болгария и Родопский массив) и Jankaea (Фессалийский Олимп) и один вид того же семейства — Ramonda nathaliае (Южная Югославия и Северная Греция). Очень близкий вид — R. serbica — распространен в Албании, Южной Югославии, Северо-
Западной Болгарии и Северо-Восточной Греции. Иллирийская провинция, особенно Албания, Северная Македония, Сербия и Родопский массив, очень богата эндемичными видами, многие из которых являются древними третичными реликтами с отдаленными географическими связями с восточноазиатскими и гималайскими видами. Наиболее интересны эндемичная ель Picea omorika (Центральная Югославия), близкая к гималайской P. spinulosa, сосны (Pinus heldreichii и Р. рейсе, последняя близка к гималайской P. griffithii), ревень (Rheum rhaponticum, Юго-Западная Болгария), конский каштан (Aesculus hippocastanum), маслинное Forsythia europaea (Северная Албания и прилегающие части Югославии), петров крест (Lathraea rhodopea, Родопский массив), очень близкий к японскому виду L. japonica, и другое непаразитное норичниковое Wulfenia baldaccii (Северная Албания), бурачниковое Solenanthus scardicus (Восточная Албания и Западная Македония), сложноцветное Amphoricarpus neu-mayeri (Албания, Македония, Черногория, Далмация и Герцеговина), лилейное Narthecium scardicum (Албания, Северная Македония, Чер-ногорье), диоскорейное Dioscorea balcanica (Северная Албания, Южная Югославия) и др.
Среди реликтовых форм имеется также ряд видов, ареал которых выходит за пределы Иллирийской провинции. Сюда относятся платан (Platanus orientalis), острия (Ostrya carpi-nifolia), самшит (Buxus sempervirens), клекачка (Staphylea pinnata), виноград (Vitis vinifera), сибирка (Sibiraea altaiensis), хурма (Dio-spyros lotus), сирень (Syringa vulgaris) и др. В растительном покрове наиболее характерны леса. В нижней зоне преобладают листопадные дубовые леса (Quercus robur, Q. pubescens, Q. cerris, Q. trojana, Q. pedunculiflora, Q. frainetto). Выше расположен пояс горных лесов, частью буковых, частью хвойных (из пихты, ели и эндемичных сосен Pinus heldreichii и Р. рейсе). Выше лесной границы распространены заросли соснового стланика из Pinus mugo с участием некоторых видов мелких рододендронов или заросли верескового кустарника Bruckenthalia spiculifolia. Далее следуют высокогорные луга.
Эвксинская провинция. Территория этой провинции охватывает Добруджу, Странджу (турецкое название Истранджа) и прилегающие районы Черноморского побережья Балканского п-ова, Северную Анатолию и Западный Кавказ (Аджария, Абхазия и район Кутаиси). На севере граница провинции проходит несколько севернее Туапсе по водораздельному хребту, отходящему от Главного Кавказского хребта. Интересно, что небольшие островки (эксклавы) эвксинскои флоры встречаются на Каздаге (Мизия) и Мураддаге (Фригия).
Уже давно указывалось на то, что сходство флоры Восточной Болгарии и приморской части Европейской Турции с Колхидой значительно больше, чем с близлежащими частями Балканского п-ова, расположенными к западу от хр. Истранджи и р. Марицы. С другой стороны, с колхидской флорой тесно связана также флора Северной Анатолии — от Босфора до Лазиста-на. На этом основании В. П. Малеевым была установлена самостоятельная Эвксинская провинция, объединяющая все эти районы флоры колхидского типа, расположенные с трех сторон Черного моря, в древности называвшегося Эв-ксинским. Флора Эвксинскои провинции, особенно лесная, в основном чисто реликтовая, сохранившаяся с третичного периода. Многие ее элементы родственны видам восточноазиат-ской и гималайской флоры, с одной стороны, и макаронезийской и европейской — с другой. В прошлом флора колхидского типа, но гораздо более богатая, покрывала значительные пространства на территории Европы и Западной Азии.
Во флоре Эвксинскои провинции имеются лишь два монотипных эндемичных рода -зонтичное Chymsidia (Северная Колхида) и бурачниковое Megacaryon (Северная Анатолия), а также почти эндемичный монотипный род Pachyphragma (крестоцветное) и общий с Иллирийской провинцией эндемичный род Amphoricarpus (сложноцветное). Эндемичных видов много (более 200), причем в большинстве случаев это древние, реликтовые виды. Из наиболее интересных эндемичных видов эвксинскои флоры отметим пихту Нордмана (Abiesnrdmanniana, Колхида и Северная Анатолия), представителей рода эпимедиум из семейства барбарисовых (Epimedium pubigerum, Восточная Болгария, Северная Анатолия и Аджария; Е. pinnatum subsp. colchicum, Колхида и Лазистан), древний реликтовый дуб с листьями, очень похожими на листья каштана (Quercus ponLica, Колхида и Лазистан), и другой реликтовый дуб (Q. hartwissiana, Колхида, бассейн р. Белой на Северном Кавказе, Северная Анатолия, Восточная Болгария и восток европейской части Турции), реликтовые примитивные березы (Веtula medwedewii, Гурия, Аджария и Лазистан, В. megrelica, известковые хребты Мегрелии), лещины (Corylus colchica, Северная Колхида; C.pontica, Колхида и Лазистан), пион (Paeonia wittmaiiniana, Колхида и Северный Лазистан), зверобои (Hypericum bupleuroides, Колхида и Лазистан; H.xylosteifolium, Колхида и Лазистан), древовидные рододендроны (Rhododendron ungernii и R. smirnovii, Колхида и Лазистан), очень редкий реликтовый кустарник из вересковых Epigaea gaultherioides, первоцвет (Primula megaseifolia, Аджария и Лазистан), цикламен (Cyclamen adsharicum, Аджария и Лазистан), андрахне (Andrachne colchica, на известняках нижнего лесного пояса от южной части Аджарского района до Кутаиси), болотный орех (Trapa colchica, Колхида), кле-качку (Staphylea colchica, Колхида), колхидский плющ (Hedera colchica, Колхида и Лазистан, на востоке в качестве реликта встречается в лесах Боржомского района), зонтичные (Astrantia pontica, Северная Колхида; Heraceum mante-gazzianum, Колхида с иррадиациями; Seseli rupicola, Адлерский район и Абхазия к северу от Кодера), крушину (Rhamnus imeretinus, Колхида и Лазистан), османтус (Osmanthus decorus, Аджария и Лазистан), бурачниковое Trachystemon orientalis (Колхида, Северная Анатолия, Восточная Болгария и Фракия) и несколько видов бурачникового рода Ompha-lodes, норичниковое Rhamphicarpa medwedewii (Колхида) и очень оригинальную горечавку (Gentiana paradoxa, известняки Колхиды), ворсянковое Scabiosa olgae (небольшой район от Туапсе до Бзыби) и несколько других видов этого рода, колокольчики (Campanula mirabilis, известняки Колхиды, и С. lactiflora, Колхида и Лазистан), девясил (Inula magnifica, Колхида и Лазистан), лилию (Lilium ponticum, Аджария и Лазистан), иглицу (Ruscus colchi-cus, Колхида, Северная Анатолия и южная часть Истранджи), диоскорею (Dioscorea caucasica, Колхида), несколько видов осоки (Сагех) и др.
Многие эндемичные виды, такие, как парадоксальная генциана (Gentiana paradoxa) и удивительный колокольчик (Campanula mirabilis), приурочены к известнякам. Некоторые колхидские элементы встречаются островками и далее на восток, например кавказская пихта (Abies nordmanniana) в Кабардино-Балкарской АССР. Господствующие растительные формации Эвксинскои провинции — леса, главным образом дубовые, буковые и буко-во-каштановые, которые выше в горах сменяются пихтовыми. Характерно наличие вечнозеленого подлеска (Rhododendron ponticum, Laurocerasus officinalis), а также таких интересных листопадных кустарников, как черника (Vaccinium arctostaphylos — вид, очень близкий к V. padifolium на о. Мадейра). Из лиан наиболее характерны колхидский плющ и сассапариль (Smilax excelsa). Выше лесов расположены высокогорные луга, а вдоль морского побережья — низинные смешанные и ольховые леса и болота. Колхидская флора лучше всего сохранилась в Аджарии и Лазистане (наибольшая концентрация лесных реликтов) и наиболее обеднена на востоке Балканского полуострова.
Кавказская провинция охватывает горные системы Большого и Малого Кавказа, но без западных частей, относящихся к Эвксинской провинции, и без засушливых юго-западных, южных и юго-восточных частей, относящихся к Армено-Иранской провинции. Сюда входят также южная предгорная часть Кубанской низменности к югу от р. Кубань, большая часть Ставропольской возвышенности, Тер-ско-Сунженская возвышенность и весь Дагестан, а также Центральное Закавказье.
Рионская низменность, Ашнеронский полуостров, Куринская и Ленкоранская низменности, долина Аракса и Талышские горы не относятся к территории Кавказской провинции.
Флора Кавказской провинции, особенно ее западной части, имеет много общего с флорой Эвксинской провинции, особенно Колхиды. Имеется целый ряд общих эндемиков, в том числе два общих эндемичных рода — зонтичное Agasyllis и первоцветное Sredinskya, кавказский рододендрон (Rhododendron caucasicum), древовидная черника (Vaccinium arctostaphylos), волчник (Daphne pontica), вороний глаз (Paris incompleta). В высокогорной флоре много общего с Пиренеями, Апеннинами, Альпами, Карпатами и Балканами (Иллирийская провинция). В Дагестане и в некоторых частях Малого Кавказа (особенно бассейн оз. Севан, Зангезур и Карабах) очень заметно выражено влияние армено-иранской и отчасти туранской флоры. Во флоре Кавказской провинции наиболее многочисленны сложноцветные, за которыми (в убывающем порядке) следуют злаки, бобовые, розоцветные, гвоздичные, крестоцветные, зонтичные и губоцветные. Имеется 5 эндемичных родов — крестоцветное Pseudovesicaria, зонтичное Symphyoloma, валериановое Pseudobetckea, бурачниковое Trigonocaryum. и сложноцветное Cladochaeta.
Видовой эндемизм довольно высокий, особенно в высокогорьях центральной части Большого Кавказа (Балкария, Дигория) ив Дагестане. Из эндемичных видов укажем, например, березу Радде (Betula raddeana), мак (Pa-paver oreophilum), хохлатки (Corydalis pallidiflora и С. emanuelii), ясколки (Cerastium kashek, С. argenteum и С. multiflorum), минуарцию (Minuartia inamoena), смолевку (Silena lacera), гипсолюбку (Gypsophila acutifolia), душистую гвоздику (Dianthus fragrans), крестоцветные (Sobolevskia caucasica, Draba bryoides, D. elisabethae, D. supranivalis, D. molissima, D. ossetica), первоцвет (Primula bayernii), камнеломку (Saxifraga subverticillata), очиток (Sedum stevenianum), герань (Geranium renardii), остролодочник (Oxytropis owerinii), целый ряд видов рода Astragalus, горечавки (Gentiana grossheimii, G. lagodechiana и G. marcowiczii), веронику (Veronica caucasica), колокольчик (Campanula andina), василек (Centaurea amblyolepis), лилию (Lilium monadelphum), подснежник (Galanthus latifolius), птицемлечник (Ornithogalum magnum), безвременник (Colchicum laetum), асфодель (Asphodeline tenuior), гусиный лук (Gagea helenae), вейник (Calamagrostis caucasica). Некоторые из эндемичных кавказских видов иррадиируют, т. е. несколько выходят за границы провинции, заходя в пограничные части соседних провинций.
Растительный покров Кавказской провинции очень пестрый. Из лесных формаций наиболее низкое положение занимают низменные прибрежные (пойменные) и болотистые леса, за которыми следуют очень характерные для провинции горные лиственные леса. Наиболее характерны леса с господством бука и скального дуба (Quercus petraea). Верхний горный пояс и отчасти средний в западной половине Большого Кавказа заменен хвойными лесами. В центральной и восточной частях северного склона Большого Кавказа нижний горный пояс занят сейчас, как правило, лесостепью, представляющей чередование дубовых насаждений с луговыми степями. Предполагают, что раньше дубовые леса занимали здесь более обширные пространства. Выше буковых лесов располагается пояс субальпийских парковых лесов, образованных в основном кленом Траутфеттера (Acer trautvetteri) и восточным дубом (Quercus macranthera). Здесь развивается березовое криво лесье, занимающее обычно более скалистые и крутые склоны. Выше располагаются альпийские луга. Степная растительность развита главным образом по невысоким хребтам в Восточном Предкавказье (Терский, Кабардино-Сунженский и др.).
Восточно-Европейская провинция. На западе в состав Восточно-Европейской провинции входят юго-восточная часть Прибалтики, Белорусская гряда, Полесье, Подольская возвышенность, южные предгорные равнины Карпат и Нижнедунайская низменность. На севере она ограничена зоной хвойных лесов Северо-Восточно-Европейской провинции (см. дальше), а на юге простирается до северного побережья Черного моря (за исключением Южного берега Крыма), северного и восточного побережий Азовского моря, долины Кубани, Ставропольской возвышенности, Волгограда и Уральска. На востоке ее граница приблизительно совпадает с восточной границей дубовых лесов и сопровождающего их «неморального комплекса», которые в районе Южного Урала переваливают хребет. В целом северная, восточная и юго-восточная границы провинции более или менее соответствуют распространению летнего дуба (Quercus robur), платановидного клена (Acer platanoides), обыкновенного орешника (Corylus avellana), бересклета (Euonymus verrucosus), копытня (Asarum europaeum), песчаного астрагала (Astragalus arenarius), вики кашубской (Vicia cassubica), чины лесной (Lathyrus sylvestris), колокольчика персиколистного (Campanula persicifolia), ландыша (Convallaria majalis), осоки горной (Carex montana). Несмотря на обширность территории, родовой эндемизм во флоре Восточно-Европейской провинции не выражен, но в южных степных районах есть эндемичные или почти эндемичные секции и подсекции рода Centaurea (василек). Число эндемичных видов относительно небольшое. Из числа эндемичных видов укажем ветреницу (Anemone uralensis), мак (Papaver maeoticum), виды гвоздики (Dianthus eugeniae, D. Krylovianus и D. volgicus), двурядник меловой (Diplotaxis cretacea), гулявник волжский (Sisymbrium wolgense), сирению Талиева (Syrenia talievii), грушу русскую (Pyrus rossica), виды копеечника (Hedysarum cretaceum и Н. ucrainicum), журавельник Бекетова (Erodium beketowii), льнянки (Linaria cretacea и L. macroura), норичник меловой (Scrophularia cretacea).
В плейстоцене флора территории Русской равнины претерпела значительные изменения, связанные с оледенением. Более древняя флора сохранилась лишь на возвышенных местах, не подвергшихся оледенению. К числу таких реликтовых районов относятся Подольская возвышенность, Донецкий кряж, гранитные выходы в районе Жданов — Бердянск, правобережье Днепра, некоторые возвышенности в Курской, Воронежской, Харьковской и Тульской областях с выходами мела и известняков («горные боры»), приволжские возвышенности (в частности, Жигулевские горы и район Пензы) и Средний и Южный Урал.
В послеледниковое время территория Восточно-Европейской (Русской) равнины заселилась растениями из этих реликтовых районов, а также из реликтовых центров Центральноевропейской и Кавказской провинций.
В настоящее время растительный покров Восточно-Европейской провинции образован главным образом широколиственными и смешанными хвойно-широколиственными лесами (Сарматская подпровинция), а на юге - луговыми лесостепями и степями (Понтическая, или Южно-Русская, подпровинция). Смешанные хвойно-широколиственные леса занимают обширную площадь в западной части провинции (от юго-запада Ленинградской области на севере до Бреста и Брянска на юге), но лишь узкой полосой идут к востоку. Здесь преобладают еловые леса из Picea abies, но со значительной примесью широколиственных пород. Начиная с Карельского перешейка (севернее ст. Отрадное) начинает попадаться летний дуб (Quercus robur), северная граница которого идет далее к Ладожскому озеру (на островах которого он растет севернее Приозерска). По восточному берегу Ладожского озера, южнее устья Свири, граница летнего дуба через окрестности Тихвина пересекает Вологодскую (около Череповца) и Кировскую области, откуда проходит в Предуралье, в Пермскую и Свердловскую области. На южной оконечности Урала дуб переваливает невысокие здесь горы и уходит сравнительно далеко за пределы Восточно-Европейской провинции. Зона дубовых лесов простирается от юго-запада к северо-востоку, образуя два широких выступа к югу, в пределах Приволжской возвышенности и Предуралья. В этих лесах встречаются ясень (Fraxinus excelsior), клены, груша, пязы, липа (Tilia cordata), ольха (Alnus glutinosa) и др. Из кленов наиболее широко распространен платановидный, или остролистный, клен (Acer platanoides), который вслед за дубом заходит за Южным Уралом на Салаирский кряж (на севере он переходит на восточный склон Урала близ Уфы). Полевой клен (Acer campestre) также входит в состав дубовых лесов, но только северная граница его проходит значительно южнее, с широты Чернигова по направлению к Орлу, Туле, откуда в Тамбовскую и Саратовскую области, но берегов Волги он не достигает. Подобно полевому клену, распространен и татарский клен (Acer tataricum), восточная граница которого достигает г. Оренбурга. Обыкновенная груша (Pyrus communis) доходит на восток почти до Волги. Все эти породы в северной части своего распространения представляют примесь к еловым лесам и входят в состав смешанных лесов. Южнее они образуют подзону широколиственных лесов, в составе которой в западной части Белоруссии и в Приднестровье, кроме летнего дуба, встречается также скальный дуб. В Приднестровье, а также к западу от Белостока, между Бугом и Наревом, имеются дубово-грабовые леса. Во всех подзонах местами имеют значительное распространение сосновые леса. Интересно, что в Волынском Полесье, а также в Мозырском районе Белоруссии произрастает желтоцветный рододендрон из группы листопадных видов (Rhododendron luteum). На западе доходит до Югославии. Предполагают, что это третичный реликт, основной ареал которого в настоящее время находится в Западном Закавказье и Северной Анатолии. В южных частях Восточно-Европейской (Русской) равнины местами большие пространства занимает лесостепье, в котором участки лесной растительности чередуются с луговыми или разнотравными степями. Преобладающей лесной породой лесных участков лесостепи здесь обычно является летний дуб, но в юго-восточной части это господство местами переходит к березе. Березовые рощи характерны для Кунгурского и Красноуфимского лесостепья, а также для лесостепья к югу от Белебея и местами в Заволжье. Из хвойных пород в лесах лесостепья встречается сосна, растущая главным образом на песках и супесях, а изредка также на меловых обнажениях и гранитных скалах (например, на Кокчетавской возвышенности). Кое-где она попадается на торфяниках. Южнее простирается зона степной растительности. Степи двух типов — ковыльно-разнотравные и ковыльно-типчаковые. В прошлом большая часть территории степей, особенно более северных районов, была покрыта широколиственными лесами и лесостепьем, о чем, в частности, свидетельствуют островные местонахождения отдельных древесных пород, как, например, граба.
Северо-Восточно-Европейская провинция простирается от Норвегии до Тиманского кряжа и Верхнекамской возвышенности. Она полностью заключена в зону хвойных лесов, или тайги, которая на востоке постепенно приближается по своему флористическому составу к западносибирской тайге. Эндемичных видов и даже подвидов в этой провинции немного, и флора здесь очень молодая по своему возрасту. Из эндемичных таксонов отметим василистник (Thalictrum minus subsp. kemense), верблюдку (Corispermum algidum), кастилею (Castilleja schrenkii). Главной лесообразующей породой является обыкновенная ель с двумя ее подвидами— Picea abies subsp. abies и Р. abies subsp. obovata; причем вторая, в основном сибирская, раса становится более обычной в восточной части провинции. К ели часто примешивается обыкновенная сосна, которая нередко на песчаных местах образует чистые насаждения, занимающие иногда большие пространства. К ели и сосне постоянно примешиваются береза и осина. На востоке появляются сибирская пихта (Abies sibirica, восточнее р. Ваги) и сибирская лиственница (Larix sibirica, к востоку от Онежского озера). Им сопутствуют некоторые сибирские элементы, как ломонос (Clematis alpina subsp. sibirica), красный воронец (Actaea erythrocarpa), пион (Paeo-nia anomala) и др. В качестве подлеска в севе-ро-восточноевропейской тайге обычны рябина, виды шиповника и др.
Западно-Сибирская провинция. На восток от Северо-Восточно-Европейской и Восточно-Европейской провинций простирается обширная Западно-Сибирская провинция, доходящая на востоке до Енисея. На севере она граничит с Арктической провинцией, а на юге — с полупустынями Казахстана. На юго-востоке граница западносибирской флоры доходит до Томска и проходит несколько восточнее Новосибирска, поднимаясь вверх по долине Оби приблизительно до Бийска, а затем поворачиваем на юго-восток и далее на юг, доходя до 50° с. ш. к западу от Усть-Каменогорска. Самыми крупными семействами западносибирской флоры являются сложноцветные, бобовые, злаки, осоковые, розоцветные, гвоздичные, норичниковые, лютиковые, губоцветные.
Несмотря на обширность территории, видовой эндемизм здесь невысокий, а родовой эндемизм полностью отсутствует. Для Западно-Сибирской провинции характерно отсутствие дуба, ясеня, клена, орешника и многих других широколиственных древесных пород. Только липа сердцевидная (Tilia cordata) встречается в лесах Тобольского Севера, Нарыма и достигает на востоке Красноярска. Древовидные ольхи сменяются здесь кустарниковой ольхой (Alnus viridis subsp. fruticosa).
Большая часть Западно-Сибирской провинции занята темнохвойной тайгой. Лесообразующей породой является здесь обычно сибирская ель (Picea abies subsp. obovata), вместе с которой растут сибирская пихта и сибирская кедровая сосна (Pinus sibirica). Сибирская лиственница и сосна, особенно на песчаной почве, также играют большую роль в составе западносибирских лесов. Из лиственных пород в этих лесах обычны березы и осина. На севере тайга переходит в предтундровые редколесья и лесотундру, а на юге — в лесостепи и степь. Интересно, что флора лесостепи носит смешанный, европейско-сибирский характер. Так, в Барабинской лесостепи более 75% видов общих с Уфимской ж Саратовской лесостепью. В прошлом, однако, здесь, несомненно, преобладал сибирский элемент. Но и теперь, вследствие более суровых зим, основной древесной породой западносибирской лесостепи является не дуб, как обычно в европейской лесостепи, а береза, встречающаяся здесь небольшими изолированными рощицами, разбросанными среди безлесных участков. Самую южную окраину Западно-Сибирской низменности занимают степи, отличающиеся от европейских степей большей сухостью и менее продолжительным вегетационным периодом. Различают две подзоны — северную, или подзону ковыльно-разнотравных черноземных степей, и южную, или подзону ковыльно-типчаковых каштановых степей.
Алтае-Саянская провинция охватывает Са-лаирский кряж и Кузнецкий Алатау, Горный: Алтай с частью Северо-Западной Монголии, Горную Шорию, Минусинскую котловину, Западный и Восточный Саян, хребет Хамар-Дабан, горные склоны, окружающие южную часть оз. Байкал южнее устья р. Баргузин и междуречье Селенга — Чикой.
Во флоре провинции один эндемичный род Microstigma и более 120 эндемичных видов, особенно в родах остролодочник (Oxytropis) и астрагал (Astragalus). Из наиболее интересных эндемиков отметим каллиантемум (Callianthe-mum sajanense), весенник (Eranthis sibirica), водосбор (Aquilegia borodinii), живокости (Delphinium mirabile и D. inconspicuum), акониты (Aconitum krylovii и A. altaicum), ветреницу байкальскую (Anemone baicalen-sis), лютик (Ranunculus sajanensis), травянистое барбарисовое Gymnospermium altaicum, березу (Betula kelleriaria), звездчатки (Stellaria martjanovii, S. imbricata и S. irrigua), смолевку (Silene turgida), крестоцветные (Aphragmus involucratus, Erysimum inense), молочаи (Euphorbia alpina, E. altaica и Е. tshuiensis), толстянковые (Rhodiola algida и Sedum populifolium), селезеночник (Chrysosplenium filipes), карагану (Caragana altaica), горошек (Vicia lilacina), чины (Lathyrus frolovii и L. krylovii), лен (Linum violascens), норичник (Scrophularia altaica), схизонепету (Schizonepeta annua), валериану (Valeriana petrophila) и некоторые другие виды этого рода, сложноцветные (Brachanthemum baranovii, Echinops humilis, Saussurea serratuloides, S. sajanensis), лук (Allium pumilum), осоку (Carex tatjanae).
Растительный покров характеризуется отчетливо выраженной вертикальной зональностью. Господствующими формациями являются лиственничные леса из сибирской лиственницы (Larix sibirica) и пихтовые леса из Abies sibirica, которые высоко в горах сменяются альпийскими лугами. Пихтовые леса распространены в сред-негорном поясе Алтая, Кузнецкого Алатау, Салаира, Хамар-Дабана, а в менее типичном виде в Западном Саяне.
В составе травяного покрова пихтовых (и вообще черневых) лесов наряду с обычными таежными видами присутствует ряд т. н. «неморальных» видов, т. е. видов, обычно являющихся спутниками широколиственных лесов Европейской части СССР или Дальнего Востока. К таким растениям относятся из папоротников щитовник мужской (Dryopteris filixmas), из цветковых растений копытень европейский (Asarum europaeum), зонтичное Osmorhiza aristata (основной ареал на Дальнем Востоке), подлесник (Sanicula europaea), кипрей горный (Epilobium montanum), двулепест-ник парижский (Circaea lutetiana), ясменник (Asperula adorata), колокольчик (Campanula trachelium), овсяница гигантская (Festuca gigantea). Наиболее богаты такими реликтами черневые леса Кузнецкого Алатау.
Часто наблюдается значительная примесь к пихте осины. Нередко присутствует сибирская кедровая сосна, реже ель и береза. В некоторых местах Кузнецкого Алатау, в истоках р. Томь, в составе темнохвойных лесов встречается липа, которая на небольшой площади образует почти чистое насаждение (Кузедеевский «липовый остров»). Для низкогорных и предгорных районов характерны парковые светлые леса из сибирской лиственницы с богатым травяным покровом. В подлеске наряду с другими кустарниками встречается даурский рододендрон (Rhododendron dauricum). Местами довольно типичны сосновые леса из Pinus sylvestris.
В предгорьях и межгорных котловинах характерны островные степи, площадь которых увеличивается к югу. В степях, помимо злаков и разнотравья, во многих местах видную роль играют кустарнички, в том числе карагана и бобовник (Prunus tenella). В горах произрастает эдельвейс (Leontopodium ochroleucum).
Среднесибирская провинция. Эта провинция охватывает обширную часть Северной Сибири, от нижнего и среднего течения Енисея, междуречья верхнего течения Ангары и Лены и устья Баргузина (впадает в оз. Байкал) до Верхоянского хребта и правобережья среднего течения Алдана.
В то время как в Западной Сибири преобладают древесные породы алтайского центра послеледникового расселения, т. е. ель, сибирская пихта, сибирская кедровая сосна и сибирская лиственница, в Среднесибирской провинции заметно участие древесных пород забайкальского и отчасти маньчжурского центров, в том числе даурской лиственницы Larix gmelinii (L. dahurica), представляющей собой основную лесообразующую породу.
Во флористическом отношении границы провинции очень смазаны. Для нее, особенно для западной ее части, характерно, что по ее территории проходит множество границ западных видов. Так, например, по Енисею проходит восточная граница Corydalis halleri, по Бирюсе - восточная граница Lathyrus vernus, по Ангаре — восточная граница смолевки Silene otites и т. д.
Во флоре Среднесибирской провинции нет эндемичных родов, а число эндемичных видов и подвидов невелико.
По Л.. В. Шумиловой (1962), для Средней Сибири характерно почти меридиональное простирание горизонтальных зон к северу от 60-й параллели: от темнохвойной тайги близ Енисея к лиственничным лесам восточнее и предтундровым редколесьям на Енисейско-Ленском водоразделе, а также наиболее полное для всего материка Евразии развитие зоны сосновых лесов на юге.
В Средней Сибири наряду с северными редкостойными чистыми лиственничными лесами, иногда с примесью березы (Betula platyphylla), характерны распространенные южнее редкостойные заболоченные лиственничные леса с участием сибирской ели и обыкновенной сосны (Pirius sylvestris). В долинах рек встречаются небольшие насаждения сибирской ели. Далее на юг, примерно до 65° с. ш., тянется зона лиственничных лесов с примесью всех предыдущих пород и, кроме того, сибирской кедровой сосны и сибирской пихты. Еще далее к югу, примерно до 60° с. ш., идет зона темно-хвойно-лиственничных лесов. Наряду с господствующей здесь даурской лиственницей характерны ель и часто кедровая сосна, а также пихта, которая образует даже чистые насаждения, тянущиеся на десятки километров вдоль рек.
На склонах водораздельных возвышенностей, покрытых кедрово-елово-лиственничной тайгой, хорошо развит подлесок из ив, кустарниковой ольхи, шиповника иглистого (Rosa acicularis), сибирской рябины (Sorbus aucuparia subsp. sibirica). В травяном покрове характерен борщевик. В этой зоне имеются также елово-пихтовые леса, для которых характерна линнея северная (Linnaea borealis, семейство жимолостных). Еще южнее, преимущественно в бассейне Ангары, а также в верховьях Нижней и Подкаменной Тунгусок и в верховьях Лены до Байкальского хребта, распространены сосновые леса, часто с подлеском из даурского рододендрона. В бассейне верхнего течения Лены широко распространены елово-листвен-ничные леса. В южной части зоны к ним присоединяются участки лесостепей. К югу распространены лиственнично-сосновые леса, но лиственница в юго-западной части провинции сибирская. Лиственничные леса в восточной части, особенно на песках, имеют примесь сосны с подлеском из сибирской рябины, ивы, кустарников ольхи и с богатым травяным покровом.
Имеются степные и лугово-степные участки, приуроченные к поймам и террасам, водоразделам и озерным котловинам. Здесь доминирующую роль играют типичные степняки — типчак, полыни, ковыль, тонконог и др.
Забайкальская провинция занимает территорию, простирающуюся на восток и юго-восток от Патомского и Северо-Байкальского нагорий и включающую северную часть Прибайкалья и большую часть Забайкалья, а также часть Северо-Восточной Монголии. Имеется один близкий к роду Раггуа монотипный эндемичный род Borodinia из семейства крестоцветных, встречающийся в южной части Байкальского хребта и почти на той же широте на Баргузин-ском хребте, а также на хребте Кодар. Несмотря на обширность территории, эндемичных видов и подвидов немного. Из эндемичных видов отметим борца (Aconitum montibaicalensis), крупку (Draba baicalensis), камнеломку (Saxifraga algisii), лапчатку (Potentilla adenotricha), астрагала (Astragalus trigonocarpus), остролодочника (Oxytropis heterotricha), мертензию (Mertensia serrulata, бурачниковое).
Господствующая растительность Забайкальской провинции представлена горными лиственничными лесами, причем лиственница представлена почти всюду здесь Larix gmelinii, за исключением юго-западной части провинции, где произрастает Larix sibirica. На склонах долин к даурской лиственнице изредка примешиваются сибирская сосна и пихта сибирская. На более сухих южных склонах встречаются сосновые боры из обыкновенной сосны. Выше границы леса располагается пояс кедрового стланика, тощей березы (Betula nana subsp. exilis), березки (Betula divaricata) и кустарниковой ольхи. Еще выше начинается горная тундра.
Для Забайкальской провинции, особенно южных ее частей, характерны острова лугово-степной, степной и лесостепной растительности, приуроченные к межгорным котловинам. Самая северная — Баргузинская степь. Наряду со степями, в которых господствуют такие виды злаков, как леймус (Leymus chinensis), житняк, типчак, ковыль, тонконог и др., и злаковыми степями с участием видов караганы (Caragana, семейство бобовых) характерны так называемые разнотравные степи, в которых преобладают двудольные растения. В разнотравных степях часто встречается красивое лилейное растение Hemerocallis minor, которое иногда даже господствует, и в таких случаях разнотравная степь переходит в так называемую желтолилейную степь.
На флору луговых степей, степей и лесостепей заметное влияние оказывает восточноазиат-ская флора, причем это влияние возрастает по направлению к востоку.
Алдано-3ейская провинция охватывает бассейн Алдана и большую часть бассейнов Зеи, Селемджи и Бурей. Флора этой провинции значительно беднее, чем Забайкальской, а растительность менее разнообразна. Основу растительного покрова составляют различные лиственничные леса, образованные даурской лиственницей. Помимо лиственничных лесов, характерны сосново-лиственничные и сосновые леса из сосны обыкновенной. В горном Юдомо-Майском районе довольно часто встречаются белоберезники. Встречаются долинные леса из ели сибирской, а местами, у верхней границы леса в Юдомо-Майском районе, в верховьях Зеи, на Становом хребте, появляются леса из ели аянской (Picea jezoensis). У верхней границы леса встречаются также рощи каменной березы (Betula ermanii). Широко распространены различные болота, заросли кустарниковой или ерниковой березки (Betula fruticosa) и сырые злаковые луга с вейником Лангсдорфа (Calamagrostis langsdorfii). Местами заметную роль играют группировки кедрового стланика и горные тундры.
Северо-Восточно-Сибирская провинция занимает обширное пространство Северо-Восточной Сибири, от западных и юго-западных предгорий Верхоянского хребта до Охотского и Берингова морей, за исключением Камчатки и областей, относящихся к Арктической провинции.
Имеется эндемичный монотипный род Gorodkovia из семейства крестоцветных и много эндемичных видов и подвидов, из которых отметим хохлатку Городкова (Corydalis gorodkovii), проломник Городкова (Androsace gorodkovii), камнеломку анадырскую (Saxifraga anadyrensis), виды лапчатки (Potentilla anadyrensis и Р. tollii), овсец (Helictotrichon krylovii), мятлик (Роа lanatiflora).
На побережье Охотского моря (п-ов Пьяги-па и южнее) встречается значительное число охотско-камчатских элементов. Единственными представителями семейства сосновых в Северо-Восточно-Сибирской провинции являются кедровый стланик (Pinus sibirica subsp. pumila) и лиственница Каяндера (Larix gmelinii subsp. cajanderi). На большей части территории провинции господствуют редкостойные и невысокие (до 10—15 м высоты) лиственничные леса, верхняя граница которых обычно лежит на высоте всего 600—700 м над уровнем моря, а в северных горных районах даже на высоте 200—400 м, и лишь в менее холодных, южных областях они изредка достигают 1100—1400 м.
В подлеске лиственничных лесов часто произрастают низкорослые березки (Betula rotundifolia, В. divaricata и В. fruticosa), виды ивовых, кустарниковая ольха, карликовый можжевельник (Juniperus communis subsp. nana) и рододендроны (Rhododendron parvifolium и R. adamsii). От редкостойных лесов каменистых горных склонов отличаются долинные леса, состоящие преимущественно из душистого тополя (Populus suaveolens) и ивы чозении (Chosenia arbutifolia). Выше лесной зоны характерны густые заросли кедрового стланика или кустарниковой ольхи, постепенно переходящие в горную тундру, среди которой встречаются отдельные участки осоково-злаковых альпийских лугов.
Охотско-Камчатская провинция включает Камчатку (без лесотундровых районов), о. Ка-рагинский, южную часть западного побережья Охотского моря и Татарского пролива на юг приблизительно до бухты Советская Гавань, низовья Амура, весь бассейн Амгуни, Северный Сахалин на юг примерно до 51°30' с. ш., Курилы к северу от «линии Миябе», проходящей между островами Итуруп и Уруп, Командорские о-ва и западные и средние Алеутские о-ва приблизительно до 165° з. д. на востоке.
Наиболее характерными древесными породами этой провинции является пихта изящная (Abies gracilis, камчатский эндемик), пихта почкочешуйчатая (Abies nephrolepis, распространена только на материковой части провинции, где заходит на севере до южного побережья Охотского моря, а на западе до бассейна Зеи; распространена также в Приморье, Северо-Восточном Китае и в северной части п-ова Корея), аянская ель, княжик (Clematis ocho-tensis, небольшая лиана из семейства лютиковых), каменная береза, кустарниковая береза, ольха камчатская (Alnus kamtschatica), осина Давида (Populus tremula subsp. davidiiana), тополь душистый, ива чозения.
Флора нижнего Амура имеет много маньчжурских элементов, т. е. представителей восточно-азиатской флоры, о которой; речь пойдет дальше. К северу влияние маньчжурской флоры постепенно угасает. На севере флора Охотии содержит много общих элементов с Камчаткой и Якутией. С другой стороны, флора Камчатки и особенно Командорских о-вов богата берингийско-североамериканскими элементами. Имеется один монотипный эндемичный род Redowskia (крестоцветное) и довольно много эндемичных видов, в том числе изящная пихта, камчатская ель (Picea kamtschatkensis), охотская живокость (Delphinium ochotense), виды аконита (Aconitum ajanense, A. ochotense), хохлатка Редовского (Corydalis redowskii), звездчатка (Stellaria peduncularis), песчанки (Arenaria redowskii, Lychnis ajanensis), камчатская рябина (Sorbus kamtschatcensis), остролодочник (Охуtropis ajanensis, 0. tilingii), камчатская бузина (Sambucus kamtschatica).
Канадская провинция включает большую часть Аляски и Канады (в том числе большую часть Лабрадора) за исключением тундры. Общий характер растительного мира Канадской провинции очень напоминает восточносибирскую тайгу, но леса отличаются здесь большим богатством лиственных пород. Наиболее распространенными породами являются виды ели (Picea mariana и P. glauca, последняя очень близка к обыкновенной ели P. abies), лиственница тамарак (Larix laricina), близкая к Larix gmelinii, осина (Populus tremuloides), близкая к P. tremula, береза (Betula papyrifera), близкая к евросибирской В. pubescens. В юго-восточной части провинции встречаются американская туя (Thuja occidentalis), канадская цуга (Tsuga canadensis), белая сосна (Pinus strobus) и красная сосна (Pinus resinosa), что связано с влиянием на флору Канадской провинции флоры Атлантическо-Северо-Американской области.
Характерными кустарниками Канадской провинции являются можжевельники (Juniperus communis и J. horizontalis), виды рода Ribes (смородина и крыжовник), кустарниковая лапчатка (Potentilla fruticosa — вид, распространенный также в Европе, Сибири и Восточной Азии), крушина (Rhamnus alnifolius), шеффер-дия (Shepherdia canadensis) из семейства лоховых, кизил (Cornus stolonifera), близкий к евразиатскому С. alba, и ряд представителей семейства жимолостных.
В травяном покрове много видов, общих с евросибирскими или близких к ним.
2. Восточно-Азиатская (Японо-Китайская область).
В эту область входят Восточные Гималаи к востоку от Западного Непала, Верхний Ассам, Северная горная Бирма, Северный горный Тонкий (ДРВ), континентальный Китай за исключением тропических частей, Тайвань, п-ов Корея, Рюкю, Каюсю, Сикоку, Хонсю, Хоккайдо, Бонин и Волкано, южные острова Курильской гряды к югу от так называемой «линии Миябе», Южный и Центральный Сахалин к югу от 51°30' с. ш., Приморье и значительная часть бассейна Амура, а также юго-восточная часть Забайкалья и участки на северо-востоке и востоке Монголии.
Флора Восточно-Азиатской области чрезвычайно богата и своеобразна и насчитывает около 20 эндемичных семейств и более 300 эндемичных родов, не говоря об очень большом числе эндемичных видов.
Из эндемичных семейств Восточно-Азиатской области отметим только древнее голосеменное семейство гинкговых (Ginkgoaceae), хвойные семейства головчатотисовых (Cephalotaxaceae) и сциадопитисовых (Sciadopityaceae), довольно примитивные цветковые семейства троходендровые (Trochodendraceae), тетрацентровые (Tetracentraceae) и церцидифилловые (Cercidiphyllaceae) и семейства цирцеастровые (Circaeasteraceae), эвкомиевые (Eucommiaceae) и давидиевые (Davidiaceae). Из эндемичных родов можно упомянуть хвойные Amentotaxus, Pseudotaxus, Keteleeria, Pseudolarix, Cathaya, Metasequoia, Cryptomeria и Microbiota, акебию (Akebia) из семейства лардизабаловых, роды Kingdonia и Megaleranthis (Корейский п-ов) из лютиковых, роды Hylomecon, Eomecon и Маcleaya из маковых, роды Disanthus, Loropetalum, Corylopsis, Fortunearia и Sinowilsonia из семейства гамамелидовых, род Pteroceltis из семейства ильмовых, род Idesia из семейства флакуртиевых, роды Bolbostemma и Schizopeppon из семейства тыквенных, род Clematoclethra из семейства актинидиевых, род Bryanthus из вересковых, род Schizocodon из Diapensiaceae, poды Stephanandra, Rhodotypos, Kerria, Chaenomeles, Rhaphiolepis, Prinsepia, Dichotomanthes из семейства розоцветных, род Kirengeshoma из гортензиевых, род Tanakea из камнеломковых, род Maakia из бобовых, роды бархат (Phellodendron), понцирус (Poncirus) и псилопеганум (Psilopeganum) из рутовых, род Таpiscia из стафилейных, род Dipteronia из кленовых, роды Fatsia, Tetrapanax, Diplopanax, Evodiopanax и Kalopanax из аралиевых, род Hovenia из крушинных, роды Dipelta и Kolkwitzia из жимолостных, род Oreocharis из геснериевых, род Paulownia из бигнониевых, роды Ombrocharis, Paralamium и Perillula из губоцветных, роды Popoviocodonia, Platycodon и Hanabusaya из колокольчиковых, роды Callistephus, Parasenecio и Symphyllocarpus из сложноцветных, роды Chionographis, Metanarthecium, Heloniopsis, Tricyrtis, Cardiocrinum, Hosta, Reineckea, Nomocharis, Milula, Rohdea, Liriope, Aspidistra и Lycoris из лилейных, бамбучок (Sasa), бамбук филлостахис (Phyllostachys) и роды Oreocalamus, Shibataea, Phaenosperma и Chikusichloa из злаков, пальму Trachycarpus, ароидное Pinellia и др. В этот список мы не включаем роды монотипных семейств, упомянутых выше.
Флора Восточно-Азиатской области представляет исключительный интерес как с точки зрения истории развития умеренных флор северного полушария, так и для решения многих вопросов эволюции и систематики цветковых растений. Эта область является одновременно одним из главных центров развития высших растений, особенно голосеменных и цветковых, и одним из центров сохранения древних форм, своеобразным убежищем живых ископаемых, таких, как Ginkgo biloba. Многие роды и даже семейства, которые хорошо разграничены во флоре Циркумбореальной области, здесь связаны переходами и часто сливаются.
Восточно-Азиатская область — один из центров происхождения культурных растений и неисчерпаемый источник декоративных садово-парковых растений.
Область подразделяется на ряд провинций, из которых особый интерес представляют для нас две северные провинции — Маньчжурская и Хоккайдо-Сахалинская.
Маньчжурская (Амурская) провинция охватывает юго-восточную часть Сибири по течению Амура, Приморье (система хребтов Сихотэ-Алиня и расположенный к западу от нее район оз. Ханка, простирающийся на юг до Амурского залива), северную часть п-ова Корея бассейн среднего и верхнего течения Ялунцзяна в северо-восточной части Китая, юго-восточную часть Онон-Аргунской степи в Забайкалье и участки в Северо-Восточной Монголии (в бассейне Улдзы и Онона) и на крайнем востоке Монголии в предгорьях Хингана. В Китае западная граница огибает с запада Большой Хинган, проходит к востоку от Шэньяна (Мукдена), далее идет дугообразно в юго-восточном направлении, пересекает Ялунцзян между 40° и 41° с.ш. и образует язык, вдающийся в северную часть п-ова Корея, приблизительно до широты Пхеньяна. Отсюда граница поворачивает на север, доходит до 40° с. ш. и, проходя по хр. Пуч-жоллен, доходит до Японского моря несколько севернее мыса Орандан. На территории Советского Союза граница Маньчжурской провинции проходит приблизительно по линии переплетения границ таких восточноазиатских (в том числе маньчжурских) видов, как Pinus koraiensis, Schisandra chinensis, Corylus heterophylla, Juglans mandshurica, Actinidia kolomikta, Phellodendron amurense, Acanthopanax senticosus, Vitis amurensis, Fraxinus mandshurica.
Наиболее характерными растительными формациями Маньчжурской провинции являются широколиственные и хвойно-широколиственные леса, чем растительность этой провинции резко отличается от соседних областей Сибири. На Дальнем Востоке СССР преобладают кедрово-широколиственные леса, сочетающиеся в северных районах и горах с елово-широколиственными, а в Южном Приморье — с чернопих-тово-широколиственными лесами. Из хвойных пород наиболее обычны даурская лиственница, аянская и корейская ели, пихта почкочешуйная и пихта цельнолистная. Широколиственные леса представлены в основном дубовыми (дуб монгольский — Quercus mongolica), а в долинах горных рек ильмово-ясеневыми лесами. Из широколиственных пород, входящих в состав лесов, кроме монгольского дуба (который, несмотря на свое название, в Монголии не встречается), отметим также ясень маньчжурский (Fraxinus mandshurica), граб сердцелистный (Carpinus cordata), орех маньчжурский, грушу уссурийскую (Pyrus ussuriensis), различные виды клена, особенно клен моно (Acer mono), клен гиннала (A. ginnala), клен желтый (A. ukurunduense), клен ложно-Зибольдов, клен зеленокорый, клен маньчжурский, чубушник тонколистный, из семейства аралиевых заманиху высокую, акантопанакс тернистый (Acanthopanax senticosus), калопанакс семилопастный (Kalopanax septemlobus) и аралию высокую (Aralia elata), а также такие породы, как маакия (Maackia amurensis), дей-ция амурская и др.
Наряду с лесной растительностью видную роль в сложении растительного покрова Маньчжурской провинции играют лесостепи и различные степные группировки и лугостепи. Они характерны для юго-восточной части Забайкалья (Ононская и Цугольская степи и бассейны рек Турчи, Борзи и Урулюнгуй и от Верхней Борзи до села Аргунска), северовосточной оконечности Монголии (Ульдзинский округ) и значительных пространств в Северо-Восточном Китае. Два небольших изолированных участка встречаются в пределах советского Дальнего Востока — на Приханкай-ской равнине (бассейн оз. Ханка и средняя часть бассейна р. Суйфуна) и на Зее-Буреин-ской равнине. Характерна различного типа степная растительность, особенно степи с участием или преобладанием сложноцветного растения пижмы сибирской (Tanacetum sibi-ricum). Очень характерны также такие степные виды, как ломонос (Clematis hexapetala), песчанка (Arenaria juncea), гречиха (Polygonum alopecuroides), пион (Paeonia lactiflora), молочай (Euphorbia pallassii), бобовые растения из рода леспедеца (Lespedeza juncea и L. davurica), астрагал (Astragalus dahuricus), зонтичное ледебуриелла (Ledebouriella divaricata), скабиоза (Scabiosa comosa), вероники (Veronica linariifolia и V. davurica), шлемник (Scutellaria baicalensis), лилейное Hemerocallis minor, лук (Allium senescens), ирис (Iris dichotoma) и злаки (Spodiopogon sibiricum, Arundinella hirta, Cleistogenes chinensis).
Для Даурии,(Под Даурией в ботанико-географической литературе понимается лесостепная и степная территории Юго-Восточного Забайкалья п Северо-Восточной Монголии.) кроме пижмовой степи, характерны лиственничные леса из Larix gmelinii, а изредка встречаются также березовые леса из Betula platyphylla и сосновые. Повсюду, особенно по крутым каменистым склонам, распространены заросли степных кустарников — ильма (Ulmus macrocarpa), дикого абрикоса (Prunus sibirica), виды таволги (Spiraea aquilegifolia, S. pubescens, S. media), виды шиповника, смородины и др.
На крайнем юго-востоке, от Верхней Борзи до села Аргунска, увеличивается роль в растительном покрове черноберезовых лесов и пионовых степей. В лесостепи обычны такие деревья и кустарники, свойственные окружающим лесам, как монгольский дуб, лещина разнолистная, даурская береза (Betula davurica), вишня (Prunus glandulosa), секуринега песпедеца (Lespedeza bicolor), бересклет Маака, крушина (Rhamnus ussuriensis), жимолость (Lonicera chrysantha) и др.
В западной части провинции на флору лесо-степей и особенно степей сильное влияние оказывают соседние флористические провинции Циркумбореальной области и Монгольская провинция Ирано-Туранской области, но по направлению к востоку флора провинции становится более типично восточноазиатской. Некоторые ботаники выделяют лесостепные и степные участки Маньчжурской провинции в самостоятельную Даурскую или Даурско-Маньч-журскую провинцию, что едва ли целесообразно.
Сахалино-Хоккайдская ( С ахалино-Курилъская) провинция. Сюда входит большая часть Сахалина к югу от 51°30" с. ш., большая часть Хоккайдо к северу от депрессии Исикари и южные острова Курильской гряды — Кунашир, Шикотан и Итуруп.
Во флоре провинции имеется один эндемичный монотипный род Miyakea, близко родственный прострелу (Pulsatilla) и встречающийся только на гольцах Восточно-Сахалинского хребта. Однако видовой эндемизм относительно невысокий, и эндемики Хоккайдо-Сахалинской провинции в большинстве случаев представляют собой молодые виды или даже подвиды. Однако среди них имеются такие хорошо обособленные виды, как сахалинская пихта (Abies sachalinensis, Сахалин, Хоккайдо и южные Курильские о-ва на север до середины Итурупа). Из эндемичных цветковых растений упомянем купальницу (Trollius pulcher, о. Ришири близ Хоккайдо), каллиантемы (Callianthemum sachalinense, Сахалин; С. miyabeanum, Хоккайдо), виды борца (Aconitum sachalinense, Сахалин и Хоккайдо; A. yezoense, Хоккайдо; A. kurilense, Шикотан; A. miyabei, Центральный Сахалин; A. neosachalinense, Южный Сахалин), виды прострела (Pulsatilla sugawarei, Сахалин; P. taraoi, Курилы; P. tatewakii, Сахалин), лютик (Ranunculus yesoensis, Хоккайдо), василистник (Thalictrum integrilobum, Хоккайдо), маки (Papaver fauriei, Ришири; P. miyabeanum, Курилы), хохлатку (Corydalis culvicalcarata, Хоккайдо), смолевку сахалинскую (Silene sachalinensis), щавель (Rumex regelii), березы (Betula tatewakiana, Хоккайдо; В. apoi-ensis, Хоккайдо), несколько видов ивы (Salix), первоцветы (Primula hidakana, Хоккайдо, пров. Хидэка; P. sachalinensis, Сахалин), землянику (Fragaria yezoensis, Хоккайдо и Курилы), лапчатку (Potentilla miyabei, Хоккайдо), смородину (Ribes latifolium, Сахалин, Хоккайдо, Курилы), очиток (Sedum furusei), астрагалы (Astragalus yamamotoi, Хоккайдо; A. japonicus, Хоккайдо и Курилы; A. sachalinensis, Сахалин), несколько видов остролодочника (Oxytropis), копеечники (Hedysarum komarovii, о. Монерон близ Сахалина и Шикотан; Н. sachalinense, Сахалин и Курилы), горечавки (Gentianasugawarae, Сахалин; G. yuparensis, Хоккайдо), бубенчик курильский (Adenophora kurilen-sis, Сахалин и Курилы); эдельвейс курильский (Leontopodium kurilense, Шикотан и Итуруп), соссюреи (Saussurea chionaphylla, Хоккайдо; S. fauriei, Хоккайдо и Курилы; S. duiensis, Сахалин; S. nupuripoensis, Сахалин), полынь илистую (Artemisia limosa, Сахалин), полынь оставшуюся (A. remosa, Сахалин); бодяк гребенчатый (Cirsium pectinellum, Сахалин, Курилы и Хоккайдо); одуванчик (Taraxacum shikotanense, Курилы), стенанциум сахалинский (Stenanthium sachalinense), гусиный лук (Gagea vagina ta, Хоккайдо и Курилы), бамбучки (Sasa amphitricha, Кунашир; S. depauperata, Кунашир и Шикотан; S. sachalinensis, Сахалин и Кунашир; S. sugawarae).
Бросается в глаза, что большинство эндемиков Сахалино-Хоккайдской провинции имеет ограниченное распространение, обычно приурочено лишь к одному острову, и часто очень узколокальное распространение даже в пределах данного острова. В растительном покрове подавляющее большинство этих эндемиков играет незначительную роль.
Флора Сахалино - Хоккайдской провинции наиболее близка к флоре Японо-Корейской провинции (особенно северных частей Хонсю), но обнаруживает много общего также с флорой Маньчжурской провинции (в частности, с флорой гор Северного Сихотэ-Алиня), а также с охотско-камчатской флорой. По А. И. Толмачеву (1959), одной из наиболее ярких особенностей растительности (и флоры) Сахалино-Хоккайдской провинции является сочетание господства темнохвойных лесов с широким развитием зарослей курильского бамбука. «В такой форме это характерное для южной половины Сахалина и для Хоккайдо сочетание не повторяется нигде на земном шаре», — пишет он.
В Восточно-Азиатской области имеется еще ряд флористических провинций, на характеристике которых нет здесь возможности останавливаться. Провинции эти следующие: Японо-Корейская (юго-западная часть Хоккайдо, Хонсю, Кюсю, Танегасима, Яку, Гото, Цусима и южная часть п-ова Корея), Токаро-Окинавская (Токара, Амами и Окинава), Волкано-Бонияскдя (Вол-кано и Бонин), Тайваньская (Тайвань и Сакисима), Северо-Китайская (на север от Ланьчжоу, северного подножия Цинь-лина и Хуанхэ до южной границы Маньчжурской провинции), Центральнокитай-с к а я (почти весь хребет Циньлин, кроме его северного подножия, часть плато Гуйчжоу и южная половина лежащей к северу от Янцзы Великой Китайской равнины на север до Шаньдунcкого п-ова и Хуанхэ и на восток до Желтого моря), Фунцзяньская (часть плато Гуйчжоу и область гор Наньлин, Фу-цзянь и Чжецзянь на восток от Восточно-Китайского моря), Нагорно-Юньнаньская (Юньнаньское нагорье и прилегающие районы Северо-Восточной горной Бирмы, северных гор Лаоса и северных гор Тонкина в ДРВ), Сычуаньская (Сычуаньские Альпы и Красный бассейн), Восточно-Тибетская (Сиканская) (Восточный Тибет), Восточно-Гималайская (Восточный Непал на запад до 83° в. д., Сикким, Бутан, Верхний Ассам).
3. Атлантическо-Севере - Американская область.
Эта область простирается от Атлантического побережья и Голубого хребта до Великих равнин США включительно и занимает большую часть центральных и восточных штатов. Она вдается местами в Южную Канаду, особенно в Южном Онтарио и долине р. Св. Лаврентия. На п-ове Флорида южной границей области является тропическая зона.
Флора Атлантическо-Северо-Американской области очень богата и характеризуется довольно высоким эндемизмом. Здесь два эндемичных семейства (Hydrastidaceae u Leitneriaceae) и около 100 эндемичных родов, в числе которых маковое (Sanguinaria), крестоцветное (Leavenworthia), розоцветные (Gillenia и Neviusia), росянковое (Dionaea), акантовое (Yeatesia), лилейное (Pleea). Целый ряд родов, как Sarracenia и Uvularia, являются общими эндемиками восточной части Канадской провинции Циркумбореальной области и Атлантическо-Северо-Американской области. Видовой эндемизм очень высокий. Среди них много третичных реликтов, концентрированных преимущественно на плато Озарк и особенно в Аппалачах, главным образом на Голубом хребте.
Уже очень давно было обращено внимание на удивительное сходство флоры Атлантического побережья США, особенно Аппалачей, с флорой Восточной Азии. Еще Цуккарини в своей «Флоре Японии» (1835) привел ряд фактов, указывающих на это сходство, но этот вопрос был разработан лишь несколько позднее, в работах американского ботаника Эса Грея, который, опираясь на теорию Дарвина, объяснил связи этих флор историческими причинами. Эти связи выражаются в наличии довольно большого числа общих родов и близких, замещающих друг друга (викарных) видов, хотя в целом флора Восточной Азии значительно богаче и содержит больше примитивных форм. Интересно, что среди общих родов имеются и такие, которые представлены всего двумя видами — по одному в каждой области, например тюльпанное дерево (Liriodendron) из семейства магнолиевых. По мнению Эса Грея, подтверждающемуся современными данными, эти факты ясно указывают на существование в прошлом связи между Азией и Северной Америкой. Хотя флористический обмен между Азией и Америкой был двусторонним, но, по современным данным, основным источником служила все же флора Восточной Азии, которая, вероятно, древнее и была всегда значительно богаче. По числу примитивных семейств и родов она значительно превосходит флору Северной Америки.
Растительный покров этой области очень разнообразен и состоит из альпийской растительности типа горной тундры, разнообразных листопадных лесов, прерий, степей и водно-болотной растительности.
4. Область Скалистых гор, или Северо-Западно-Американская область.
Эта область охватывает горные системы Западной Канады и западных штатов Северной Америки, от Аляски до Калифорнии, Невады и Нью-Мексико. На юге она расходится двумя длинными «языками», разделенными Большим Бассейном.
Флора области характеризуется одним эндемичным семейством печеночников (Gyrothyraceae). Эндемичные семейства цветковых растений здесь отсутствуют. Имеется несколько десятков эндемичных или почти эндемичных родов цветковых растений, причем более чем половина этих родов монотипные. Эндемичны род Darlingtonia из семейства саррацениевых, Van-couveria (барбарисовое), Meconella (маковое), несколько родов из семейств портулаковых и маревых, больше 10 родов семейства крестоцветных, в том числе примитивный род Stanleya, род Cladothamnus (вересковое), роды Allotropa, Hemitomes, Pityopus и Pleuricospora из вертлянициевых (монотроповых), род Sidalcea (мальвовое), род Piscaria (молочайное), роды Chamaebatiaria, Luetkea, Oemleria, Petrophytum и Purshia из розоцветных, род Whipplea (гортензиевое), роды Bolandra, Lithophragma, Peltiphyllum, Suksdorfia, Tellima и Tolmiea из камнеломковых, род Orogenia (зонтичное), роды Gymnosteris, Leptodactylon и Linant-hastrum из синюховых (полемониевых), роды Hesperochiron и Romanzoffia из воднолистниковых, род Dasynotus (бурачниковое), роды Chionophila, Cordylanthus, Synthyris и Tonella из норичниковых, роды Githopsis и Howellia из колокольчиков, роды Balsamorhiza, Blepharipappus, Chrysothamnus, Crocidium, Dimeresia, Eriophyllum, Hulsea, Luina, Raillardella и Wyethia из сложноцветных, род Scoliopus (лилейное), род Scribneria (злак) и др.
Доминирующей растительностью области являются хвойные леса. Ни одна другая флористическая область земного шара не обладает столь большим разнообразием хвойных, как Тихоокеанская Северная Америка. В северной части области распространены роскошные хвойные леса, состоящие из Pseudotsuga menziesii, Pinus ponderosa, Pinus contorta, Thuja plicata, Tsuga heterophylla, Tsuga mertensiana, Picea sitchensis и Chamaecyparis nootkatensis. Дальше на юг вдоль побережья Южного Орегона и Калифорнии произрастают леса из вечнозеленой секвойи (Sequoia sempervirens), называемой также красным деревом, а в горах — из гигантской секвойи (Sequoiadendron giganteum). В этой области произрастают такие широколиственные породы, как виды клена, березы, а южнее — дуба, но наиболее широко распространенной является осина (Populus tremuloides). Для жестколистных прибрежных лесов характерно вересковое Arbutus menziesii.
В. Древнесредиземноморское царство
Древнесредиземноморское подцарство тянется от Макаронезии на западе через все Средиземноморье, Переднюю и Среднюю Азию до Монголии на востоке. Несмотря на то что растительный покров этой обширной территории чрезвычайно разнообразен и состоит из таких столь различных экологических типов, как вечнозеленые древесные растения Макаронезии и пустынные ксерофильные кустарнички Центральной Азии, по истории развития ее флора характеризуется некоторыми общими чертами. Еще один из классиков географии растений, А. Энглер (1881) в работе о географическом распространении рода Rhus (семейство анакардиевых), ввел понятие флоры «древнего Средиземноморья», имея в виду то, что она формировалась по побережью древнего Средиземного моря — Тетиса. ВпоследствииМ. Г. Попов (1927, 1929) более детально развил идею о единой древ-несредиземноморской флоре, но, к сожалению, расширил границы этой флоры вплоть до Калифорнии и Мексики, что едва ли можно признать правильным.
Древнесредиземноморская флора развивалась как преимущественно миграционная флора, и почти вся территория высыхающего моря Тетис была ареной переселений и смешения флор совершенно разного происхождения. Она развивалась на стыке бореальных и тропических флор, но подавляющее большинство древнесредиземноморских растений имеет бореальное, в частности восточноазиатское, происхождение. Поэтому Древнесредиземноморская флора является в целом флорой бореальной. Однако видоизмененные выходцы из тропической флоры играют в ней заметную роль, особенно в Макаронезийской и Сахаро-Аравий-ской областях. Сюда относятся прежде всего представители семейств лавровых (Lauraceae) и пальмы (Phoenix canariensis, в Макаронезии; Chamaerops humilis, в Западном Средиземноморье, и род Nannorrhops, распространенный от Аравии до Северо-Западной Индии). Сюда же относятся Cytinus hypocistis и Pilostyles haussknechtii из семейства раффлезиевых (Rafflesiaceae), представители семейств Capparaceae, Loranthaceae, Santalaceae и Zygophyllaceae, макаронезийский эндемик Sideroxylon marmulano и марокканский эндемик Argania spinosa — оба из семейства сапотовых (Sapotaceae), ирано-туранские виды рода Incarvillea из семейства бигнониевых (Bignoniaceae) и др. Древнесредиземноморская флора обнаруживает ясно выраженные связи с африканской, в частности с капской, флорой. Многие ксе-рофитные и особенно геофитные элементы древнесредиземноморской флоры имеют близких родичей в Южной Африке. Так, хвойное Tetraclinis articulata (семейство кипарисовых), произрастающее в Северо-Западной Африке, Юго-Восточной Испании и на Мальте, близко к роду Widdringtonia (Тропическая и Южная Африка), а также к роду Callitris (Австралия, Тасмания и Новая Каледония).
Среди двудольных африканские связи наиболее хорошо выражены в родах Limonium, Pelargonium, Erica, Argyrolobium, Convolvulus, Stachys, Salvia, Cephalaria, Ifloga, He-lichrysum, Echinops и др. Среди однодольных эти связи особенно хорошо выражены в родах Androcymbium, Colchicum, Scilla, Urginea, Dipcadi, Ornithogalum, Romulea, Gladiolus, Iris, Gynandriris и др. Эти связи Энглер объяснял общностью происхождения от исходных тропических предков, но вероятнее всего между Средиземноморьем и Южной Африкой длительное время происходил флористический обмен, который осуществлялся через горы Восточной Африки, Эфиопию и далее через Драконовые горы, которые образуют как бы мост, соединяющий древнесредиземноморскую флору с южноафриканской. Горные хребты Северо-Восточной Африки в плиоцене продолжались до Малой Азии и Балканского п-ова, чем объясняется преобладание среди среднеземно-морских элементов флоры Африки восточносреди-земноморских таксонов над западносредиземно-морскими. Этот флористический обмен был скорее всего взаимный, хотя далеко не во всех cлучаях удается определить направление миграции.
Древнесредиземноморское подцарство подразделяется на 4 области: Макаронезийскую, Средиземноморскую, Сахаро-Аравийскую и Ира-но-Туранскую.
5. Макаронезийская область.
В Макаронезийскую область входят острова: Азорские, Мадейра, Канарские и Зеленого Мыса. Флора этой области насчитывает около 1600 видов, из которых более половины средиземноморские. Эндемичных родов сравнительно мало (около 30), причем больше половины их приурочено к Канарским островам. Однако число эндемичных видов довольно велико (около 650). Из наиболее интересных эндемичных видов отметим прежде всего лавровые (Laurus azorica, Apolloniasbarbusana, Persea indica и Ocotea foetens), сохранившиеся здесь с третичного времени, когда они или близкие к ним виды были широко распространены на территории Европы и на Кавказе (особенно много их остатков в верхнемиоценовой флоре Годердзи в Западной Грузии). Далее следуют клетра древовидная (Clethra arborea, клетровое), кустарниковый щавель (Rumex lunaria, гречишное), виды кустарниковой капусты (Sinapidendron, крестоцветное), Канарское земляничное дерево (Arbutus canariensis, вересковое), Sideroxylon marmulana (сапотовое), кустарниковые молочаи (Euphorbia regis-jubae и Е. stygiana), кактусо-видный молочай (Е. canariensis) и древовидный молочай (Е. mellifera), питоспорум (Pittosporum coriaceum, питоспоровое), падуб канарский (Ilex canariensis, падубовое), бузина мадейрская (Sambucus maderensis, жимолостное), бирючина (Viburnum rugosum, жимолостное), подорожник (Plantago arborescens), достигающий 1 м высоты, древовидный василек (Gentaurea arborea), драконово дерево (Dracaena draco), финиковая пальма (Phoenix canariensis), Канарская сосна (Pinus canariensis).
Среди эндемичных видов обращает на себя внимание большое число вторичных кустарниковых и древовидных видов родов, представленных вне Макаронезии обычно травянистыми формами. Кроме перечисленных выше, упомянем виды рода молодило (Sempervivum) и близких к нему родов из семейства толстянковых, а также виды родов синяк (Echium) из бурачниковых и осот (Sonchus) из сложноцветных. Характерно наличие тропических элементов. В растительном покрове Макаронезии наиболее замечательны вечнозеленые лавровые леса, которые, как показывают данные палеоботаники, из всех современных растительных формаций наиболее близки к миоценовым и плиоценовым лесам Европы.
6. Средиземноморская область.
Средиземноморская область приблизительно в тех ее границах, которые были намечены еще А. Гризебахом (1874) и впоследствии несколько уточнены последующими авторами, включает большую часть Пиренейского п-ова, прибрежные части Франции, Апеннинского и Балканского п-овов (включая почти всю Грецию), острова Средиземного моря, Марокко, Северный Алжир, Тунис, Северо-Западную Триполитанию, Киренаику, небольшую прибрежную полосу в Северо-Западном Египте, побережье Леванта (большая часть Палестины и Ливан), Западную Сирию, Западную и Южную Анатолию. Небольшие по размерам и обедненные островки средиземноморской флоры (так называемые «эксклавы») имеются в южной, горной части Крыма и вдоль Черноморского побережья Кавказа, особенно в Северной и Северо-Восточной Анатолии.
Средиземноморская область почти лишена эндемичных семейств, если не считать монотипного семейства Aphyllanthaccae (род Aphyllanthes, Западное Средиземноморье), обычно не выделяемого из семейства лилейных. Число эндемичных родов сравнительно невелико, но много общих эндемиков с соседними областями — Макаронезийской, Циркумбореальной, а также Сахаро-Аравийской и Ирано-Туранской. Из эндемичных родов отметим западно-средиземноморский род Tetraclinis из семейства кипарисовых, роды Rupicapnos (Западное Средиземноморье) и Ceratocapnos из семейства дымянковых, крапивное Soleirolia (острова Западного Средиземноморья), гвоздичные Ortegia (Западное Средиземноморье) и Bolanthus (Восточное Средиземноморье), крестоцветные Lycocarpus (Южная Испания), Aubrietia (от Италии до Ирана). lonopsidium (Западное Средиземноморье), Bivonaea (Сицилия), Euzomodendron (Южная Испания), Hutera (Испания), Vella (Западное Средиземноморье), Boleum (Восточная Испания), Didesmus (Восточное Средиземноморье), Morisia (Сардиния и Корсика), Guiraoa (Юго-Восточная Испания), почти эндемичный западносредиземноморскип род Sesamoides из резедовых (на севере доходит до Центральной Франции), род Malope из мальвовых, росянковое Drosophyllum (Западное Средиземноморье), бобовые Ceratonia, Chronanthus, Anagyris, Calicotome, Spartium, Hymenocarpus и Biserrula, род Argania (Марокко) из тропического семейства сапотовых, зонтичные Реtagnia (Сицилия) и Lagoecia, род Putoria из мареновых, род Fedia из валериановых, род Tremastelma (Восточное Средиземноморье) из ворсянковых, норичниковые Bellardia и Lafuentia (Западное Средиземноморье), губоцветные Rosmarinus, Preslia (Западное Средиземноморье), Dorystoechas (Юго-Западная Анатолия) и Coridothymus, колокольчиковое Trachelium, сложноцветные Santolina, Cladanthus, Staehelina, Cynara (на восток до Курдистана), Leuzea, Andryala, Rothmaleria (Южная Испания), лилейное Chioriodoxa (Восточное Средиземноморье), ирисовое Hermodactylus, злак Triplachne (Сицилия), ароидные Dracunculus (от Мадейры и Канарских о-вов до Малой Азии), Helicodiceros (Балеарские о-ва, Корсика, Сардиния), Biaruni (на восток до Западного Ирана), Arisarum (на запад до Макаронезии) и, наконец, единственную в Средиземноморской области дикорастущую пальму Chamaerops (один вид С. humilis в Западном Средиземноморье). Видовой эндемизм достигает 50%, но эндемизм здесь диффузного типа, т. е. эндемики широко разбросаны среди многих родов и видов, что указывает на очень большую роль миграций. Отметим некоторые из эндемичных или почти эндемичных видов, играющих особую роль в сложении средиземноморской растительности или представляющих интерес с историко-флористической точки зрения. Это алеппская сосна (Pinus halepensis) и пиния (Pinus pinea), кипарис (Cupressus sempervirens), который дико произрастает только на островах Эгейского моря, но широко культивируется по всему Средиземноморью, благородный лавр (Laurus nobilis), Liquidambar orientalis из семейства Altingiaceae (Юго-Западная Анатолия и Родос), вечнозеленые дубы (Quercus ilex и Q. coccifera), ладанники (Cistus incanus, С. monspeliensis и другие виды этого макаронезийско-средиземноморского, преимущественно западносреди-земноморского рода), солнцецвет иволистный (Helianthemum salicifolium) и другие виды этого в основном древнесредиземноморского рода, земляничные деревья (Arbutus unedo и A. andrachne), виды вереска (Erica), древовидный молочай (Euphorbia dendroides), виды фисташки (Pistacia lentiscus и P. terebinthus), мирт (Myrtus communis), филлирея (Phillyrea angustifolia и P. latifolia), олеандр (Nerium oleander), акант (Acanthus mollis), авраамово дерево — кустарник из вербеновых (Vitex agnus-castus). Наиболее многочисленны эндемичные виды из семейств ладанниковых, крестоцветных, бобовых, зонтичных, бурачниковых, норичниковых, губоцветных, сложноцветных, лилейных и злаков.
По сравнению с флорой Макаронезийской области средиземноморская флора содержит меньше древних, третичных реликтов и гораздо больше молодых, прогрессивных эндемиков, возникновение которых было связано с возрастающей ксерофилизацией. Как и во флоре Макаронезии, влияние африканской флоры здесь ярко выражено, особенно в ксерофильных формациях и среди луковичных растений, где часто ясно видны связи с Южной Африкой, но представителей чисто тропических семейств гораздо меньше, причем они сохранились главным образом в Западном Средиземноморье, особенно в Марокко. Ясно выражена связь с восточноазиатской флорой, восходящая к третичному времени. Чрезвычайно велико влияние на средиземноморскую флору элементов цир-кумбореальной флоры, многие из которых проникли в Средиземноморье, особенно в горные его районы, в эпохи, соответствовавшие наступлению ледника на севере Евразии. В то же время в более сухие и жаркие межледниковые эпохи многие средиземноморцы проникали на север, где до сих пор они частично сохранились в виде реликтов, например в Центральной Франции, в различных районах Швейцарии и даже в Южной Ирландии, где до сих пор произрастает земляничное дерево (Arbutus unedo).
Естественный растительный покров Средиземноморья в относительно мало тронутом виде сохранился только в некоторых горных районах. Так, в Марокко, Алжире и на Сицилии все еще сохранились леса из дикой маслины.
На равнинах и в предгорьях растительный покров или полностью уничтожен и замещен культурными растениями, или очень сильно видоизменен. Для растительности Средиземноморья наиболее характерны хвойные и вечнозеленые жестколистные леса из невысоких деревьев с короткими толстыми стволами и разнообразные формации кустарников, кустарничков и полукустарников. Характер растительности заметно изменяется с запада на восток и с севера на юг. В обоих направлениях растительность становится более ксерофильной.
Средиземноморская область подразделяется на провинции:
Юго-Западно-Средиземноморская провинция занимает Южно-Атлантическую Иберию и Северное Марокко, включая Высокий Атлас. Иберийско-Балеарская провинция охватывает Субпиренейскую Иберию, исключая сами Пиренеи, и всю остальную часть полуострова, кроме районов, относящихся к предыдущей провинции и Циркумбореаль-ной области, а также Балеарские о-ва.
Лигурийско-Тирренская провинция включает Приморский Лангедок и Прованс, побережье Лигурийского моря (Французскую и Итальянскую Ривьеру), западную часть Средней Италии, итальянское побережье Тирренского моря, весь п-ов Калабрия, острова Корсика, Сардиния, Сицилия и Мальта. Адриатическая провинция занимает Адриатическое побережье Италии на север приблизительно до района Равенны и весь п-ов Салентина, все побережье п-ова Истрия, на севере включая Триест, побережье Югославии и Албании на юг приблизительно до Влёры.
Восточно-Средиземноморска я провинция охватывает часть Южной Албании, почти всю Грецию, острова Ионического и Эгейского морей, Крит, Кипр, побережье Малой Азии и Сирии, Ливан и большую часть Палестины.
Южно-Средиземноморская провинция охватывает Алжир, Тунис, Северо-Западную Триполитанию, Киренаику и часть северо-западного побережья Египта. Крымско-Новороссийская провинция охватывает Южный Крым (узкая прибрежная полоска южного склона первой гряды Крымских гор от Севастополя до Феодосии, ограниченная с севера высокогорьями Яйлы) и северную часть Западного Закавказья от Анапы примерно до Туапсе на юге. Этот сравнительно небольшой район с его довольно обедненной средиземноморской флорой, особенно в Новороссийской подпро-винции, едва ли заслуживал выделения в отдельную провинцию, если бы не его значительная удаленность от основного ареала средиземноморской флоры. Из числа типичных средиземноморцев этой провинции назовем прежде всего земляничник (Arbutus andrachne) и ладанник (Cistus inca-nus). Земляничник является характерным растением южнокрымского маквиса — обедненного варианта распространенной в Средиземноморье растительной формации. Заслуживает упоминания так называемая судакская сосна, или сосна Станкевича, встречающаяся между Судаком и Новым Светом и в противоположной, западной оконечности Южного Крыма. Обычно она рассматривается как самостоятельный вид Pinus stankewiczii, но в действительности настолько близка к средиземноморскому виду Pinus brutia, что в лучшем случае может рассматриваться лишь как его подвид. Таково же систематическое положение так называемой пицундской сосны, произрастающей на Черноморском побережье Кавказа, на мысе Пицунда в Абхазии, а также на крутых склонах хребтов, выходящих к Черному морю в районе между Абхазией и Анапой, причем вместе с ней растет также ладанник (Cistus incanus). Пицундская сосна тоже очень близка к Pinus brutia и, если не идентична ей, представляет собой лишь подвид. Эти изолированные местонахождения средиземноморской сосны имеют реликтовый характер. В прошлом, во время сухих межледниковых эпох, средиземноморцы были, вероятно, гораздо шире распространены на Черноморском побережье Кавказа и в Малой Азии. Чрезвычайно интересно, что здесь сохранилось также паразитное средиземноморское растение из тропического семейства раффлезиевых (Cytinus hypocistis).
7. Сахаро-Аравийская область.
В эту область входит вся внетропическая часть Сахары, от Атлантического побережья до Египта (по берегу залива Сидра и восточнее Киренаики достигает Средиземного моря), Синайский п-ов, вся северная внетропическая часть Аравийского п-ова, Южная Палестина, часть Иордании, южная часть Сирийской пустыни и Нижняя Месопотамия.
Флора Сахаро-Аравийской области не очень богата (около 1500 или немного более видов) и носит до некоторой степени переходный характер, так как на севере в ее составе довольно много средиземноморских растений, на юге — суданских, а на востоке — ирано-туранских. Несмотря на это, в ней довольно много эндемичных родов, а видовой эндемизм, вероятно, не менее 25%. Из эндемичных родов можно отметить Nucularia, Fredolia и Agathophora (маревые), Muricaria, Nasturtiopsis, Zilla, Oudneya, Foleyola и Lonchophora (крестоцветные), Gymnarrhena и Lifago (сложноцветные) и др. Многие наиболее характерные эндемичные виды и роды относятся к семейству крестоцветных (особенно к Brassicinae), а также к маревым, гвоздичным (особенно Paronychieae) и сложноцветным (особенно Filagineae). Характерны также эндемичные виды рода парнолистник (Zygophyllum). В растительном покрове доминируют пустынные и полупустынные формации и редколесье.
8. Ирано-Турайская область.
В эту обширную область входят Центральная и Восточная Анатолия, большая часть Сирии, Южная и Восточная Палестина, небольшая часть Синайского п-ова, часть Иордании, северная часть Сирийской пустыни, Верхняя Месопотамия, большая часть Армянского нагорья, аридные и семиаридные районы Южного и Восточного Закавказья, Гиркания (Талыш и прилегающие районы вдоль Каспийского побережья на территории Ирана), Иранское нагорье без тропических пустынь, южные отроги Гиндукуша и южные склоны и отроги Западных Гималаев к западу от 83° в. д. и вся огромная территория от низовьев Волги и пустынь Восточного Закавказья до пустыни Гоби включительно.
В таком объеме эта область приблизительно соответствует «области степей» (Steppengebiet) А. Гризебаха, установленной им сто лет назад. Флора Ирано-Туранской области характеризуется довольно высоким родовым эндемизмом и очень высоким видовым эндемизмом (вероятно, не менее 25%). Наиболее богата флора Иранского нагорья, наиболее обеднена флора Центральной Азии. Из числа эндемичных родов с более широким ирано-туранским ареалом укажем, например, таких представителей очень характерного для области семейства маревых, как Panderia, Horaninovia, Bienertia, Girgensohnia, Halocharis, Halimocnemis, Halotis, Halanthium и Gamanthus, крестоцветное Graellsia, бобовые Ammothamnus и Ammodendron, род Acanthophyllum из гвоздичных (на севере заходит в Сибирь) и род Lallemantia из губоцветных (от Анатолии до Западных Гималаев). Очень характерны для Ирано-Туранской области роды Acantholimon (свинчатковое) и Cousinia (сложноцветное), но они, как, впрочем, и многие другие ирано-туранские элементы, заходят в Восточное Средиземноморье и не являются, таким образом, строго эндемичными.
Ирано-Туранская область подразделяется на две подобласти — Западно-Азиатскую и Центральноазиатскую, границу между которыми, однако, не всегда легко указать.
8а. Западно-Азиатская подобласть.
Западно-Азиатская подобласть охватывает всю западную часть области до Прибалхашья, Центрального Тянь-Шаня и Западных Гималаев. В нее входит флористически наиболее богатая часть области, растительный покров которой отличается огромным разнообразием. В Гиркании, Западном Тянь-Шане и в Западно-Гималайской провинции еще сохранились реликтовые участки мезофильных лесов, а во многих местах значительные пространства покрыты ксерофильными дубовыми лесами. Средиземноморский элемент также выражен гораздо сильнее, особенно во флоре дубовых лесов и редколесий. Западно-Азиатская подобласть, особенно Иранское нагорье, представляет собой основной центр формирования ирано-туранской флоры. Много эндемичных видов таких родов, как Delphinium, Silene, Acanthophyllum, Calligonum, Atraphaxis, Acantholimon, Limonium, Althaea, Alcea, Euphorbia, Haplophyllum, Alyssum, Aethionema, Erysimum, Isatis, Prunus, Onobrychis, Trigonella, Ferula, Nepeta, Phlomis, Eremostachys, Salvia, Stachys, Thymus, Scrophularia, Verbascum, Heliotropium, Convolvulus, Asperula, Galium, Achil'lea, Anthemis, Tanacetum, Artemisia, Centaurea, Cousinia, Echinops, Helichrysum, Jurinea, Scorzonera, Eremurus, Tulipa, Allium, Iris.
Армено-Иранская провинция. Обширная Армено-Иранская провинция охватывает большую часть Внутренней Анатолии и семиаридные части Армянского нагорья, засушливые районы Южного Закавказья, Зувант и хребет Копетдаг на территории СССР, большую часть Ирана, за исключением южных и юго-западных прибрежных тропических районов и районов прикаспийских реликтовых лесов, часть Афганистана и Северный Белуджистан. Провинция характеризуется богатой и своеобразной флорой с очень высоким видовым эндемизмом. Имеются также эндемичные секции и подроды, более двух десятков эндемичных родов. Особенно много эндемичных таксонов в семействах сложноцветных, зонтичных, бурачниковых, крестоцветных, маревых, бобовых, губоцветных, лилейных, свинчатковых, мареновых и норичниковых. Характерны пустынные и полупустынные формации, горные степи, часто с участием колючих подушкообразных кустарничков (особенно колючий эспарцет Оnobrychis cornuta и виды Acantholimon), сообщества ксерофильных и жестколистных кустарников и кустарничков и можжевеловые и лиственные редколесья. Во многих местах распространены ксерофильные дубовые леса.
Месопотамекая провинция охватывает часть Иордании, большую часть Сирии, часть равнин Юго-Восточной Анатолии, примыкающих к Сирийской пустыне, Северо-Западный Ирак и небольшой участок Юго-Западного Ирана к западу от г. Керманшаха.
Туранская (А рало-Каспийска я) провинция включает пустыни Восточного Закавказья, Прикаспийскую низменность от низовьев Терека через низовья Волги до Эмбы, область пустынь, протягивающихся от восточных берегов Каспийского моря и низовьев Эмбы до Црибалхашья, Мугоджары, плато Устюрт, Каракумы и Кызылкум, предгорные равнины вдоль Копетдага, Памиро-Алая, Западного Тянь-Шаня, межгорные долины Мирзагуль, Каршинскую степь и другие равнины на юге Узбекистана.
Флора этой провинции обнаруживает много общего с флорой Армено-Иранской провинции, где располагаются центры происхождения очень многих ее компонентов. Очень много общего также с центральноазиатской флорой, особенно в семействе маревых (Chenopodiaceae) -наиболее характерного семейства туранской флоры. Из эндемичных или почти эндемичных туранских родов укажем Alexandra, Rhaphido-phyton и Piptoptera из маревых и род Smirnowia из бобовых. Растительный покров провинции состоит преимущественно из чисто пустынных или степных формаций. Гирканская провинция — район реликтовых лесов Талыша, северных склонов Эльбурса и Южно-Каспийской низменности до Юго-Западного Хорасана на востоке. Леса распространены вдоль побережья Каспийского моря примерно до высоты 500—600 м над уровнем моря.
Основными лесными породами являются железняк (Parrotia persica, эндемичный монотипный род из семейства гамамелидовых) и каштанолистный дуб (Quercus castaneifolia). Этот эндемичный для Гирканской провинции третичный реликт наиболее близок к произрастающему в Алжирском Атласе Quercus afares. В лесах нижнего горного пояса в качестве ле-сообразующих пород к ним присоединяются еще дзельква (Zelkova carpinifolia) и граб (Carpinus betulus).
Туркестанская провинция охватывает горы и нагорья Средней Азии до Центрального Тянь-Шаня и Памира на востоке, большую часть Бадхыза, Карабиль, Бадах-шан, Афганский Туркестан, Паропамиз, хребет Сафедкох, Западный Гиндукуш, хребет Баба, на западе до Нуристана и долины Кабула. Это во многих отношениях переходная провинция, тесно связанная на западе с Армено-Иранской провинцией, а на востоке — с цент-ральноазиатской флорой, с одной стороны, и западногималайской — с другой. Тем не менее эндемизм этой провинции довольно высокий, и в ней имеется не только много эндемичных видов, но и довольно много эндемичных родов, например крестоцветные Iskandera, Tetracmidion и др., зонтичные Albertia, Schtschurovskia, Korshinskya, Ladyginia, Komarovia, Mogoltavia и Astomatopsis, норичниковые Spirostegia и Nataniella, колокольчиковые Суlindrocarpa и Cryptocodon, сложноцветные Lepidolopha, Polytaxis и др., лилейное Korolkowia. Из большого числа эндемичных видов укажем инкарвиллею (Incarvillea olgae, Дарваз) — представителя тропического семейства бигнониевых и экзохорду (Exochorda alberti) из семейства розанных, образующую на востоке Гиссарского хребта своеобразную кустарниковую формацию. Большая часть видов обоих родов приурочена к Восточно-Азиатской флористической области.
В предгорьях и низкогорьях значительные пространства занимают пустынные типы растительности, особенно эфемеровая растительность. Для горных склонов характерны заросли листопадных кустарников, которые в большинстве случаев представляют собой вторичные сообщества, возникшие в результате уничтожения лесной растительности. Значительные пространства в горах занимают своеобразные формации степного типа с участием крупных зонтичных, эфемеров и геофитов и редколесья или реже леса, образованные можжевельниками.
Лиственные леса встречаются лишь в виде отдельных массивов. Грецкий орех образует леса в западной части Тянь-Шаня и на Гиссарском хребте. Наряду с чистыми насаждениями ореха обычны кленово-ореховые и яблонево-ореховые леса, приуроченные к более освещенным склонам. Характерны также светлые кленовые леса паркового типа из клена Семенова и туркестанского клена. Лишь небольшие пространства занимают еловые и пихтовые леса. Еловые леса состоят из ели Шренка (Picea schrenkiana) — высокого стройного дерева, распространенного на Тянь-Шане и Памиро-Алае (Иркештам), а также в Центральноазиатской подобласти. Пихта представлена в Туркестанской провинции особой географической расой сибирской пихты, известной под названием пихты Семенова. Она не образует чистых насаждений, но растет в сочетании с елью, яблоней и грецким орехом.
Высокогорная растительность представлена лугами (главным образом на Тянь-Шане), степями, сообществами подушковидных полукустарничков. Западно-Гималайская провинция включает Нуристан, долину Кабула, расположенный к югу от нее хребет Спин-rap и южные склоны и отроги Западных Гималаев приблизительно к западу от 83° в. д., в среднем выше 1000 м над уровнем моря. Отдельные эксклавы встречаются местами на восточных склонах Сулеймановых гор.
8Б. Центральноазиатская подобласть.
Центральноазиатская подобласть охватывает обширную территорию пустынь и степей, простирающуюся от Прибалхашья и Центрального Тянь-Шаня до Большого Хингана. Она занимает в основном территорию внутреннего стока Азиатского материка и представляет собой царство своеобразных холодных пустынь, пустынных и высокогорных степей. Флора ее сравнительно бедна (едва ли более 5000 видов), но эндемизм высокий и имеется, в частности, немало эндемичных родов, в том числе систематически довольно изолированных. Из числа эндемичных родов упомянем, например, маревые Microgynoecium (Тибет), Iljinia, Sympegma и Ammopiptanthus, розоцветное Potaninia (Монголия) и парнолистниковое Tetraепа (Монголия).
Из-за сухости и очень большой разницы между крайними температурами растительный мир Центральноазиатской подобласти характеризуется угнетенностью и сравнительным однообразием. Господствуют горные степи, местами переходящие в лесостепи, высокогорные луга, но особенно различные пустынные и полупустынные формации. Местами встречаются острова еловых, елово-пихтовых, лиственничных и очень обедненных лиственных лесов.
Центральнотяньшаньская провинция охватывает центральные хребты Тянь-Шаня в пределах СССР, часть Алайского хребта, Алайскую долину, а на севере - Иссык-Кульскую котловину с прилегающими склонами; на юге граница проходит по гребню Заалайского хребта; на западе — по Ферганскому хребту и далее вдоль хребта Кекеримтау и Сусамырского до Таласского Алатау; на севере — вдоль Таласского Алатау, далее по Киргизскому хребту и Кунгей-Алатау.
Флора этой провинции, несмотря на ее размеры, содержит, вероятно, лишь немногим более 1500 видов. Но в ней имеются эндемичные роды (злак Timouria, бурачниковое Tianschaniella и др.) и много эндемичных видов. В растительном покрове доминируют степные сообщества, кобрезиевые «ковры» и осоковые болота. Некоторые авторы называют Центральный Тянь-Шань страной злаков и осок. Что же касается древесно-кустарниковой растительности, то она представлена здесь фрагментарно и занимает небольшие площади. В глубине Центрального Тянь-Шаня, особенно на Терскей-Алатау и Кунгей-Алатау, имеются отдельные, обычно небольшие, островки еловых лесов. Встречаются также лиственные насаждения, состоящие из тополя, березы, видов жимолости, изредка дикого абрикоса и др.
Джунгаро-Тянынаньская провинция включает южный склон и предгорья Монгольского и отчасти Русского Алтая, хребты Тарбагатай и Саур, Джунгарский Алатау, хребты Барлык, Майли и Джаир, Восточный Тянь-Шань, Джунгарскую Гоби, Зайсанскую котловину, Джаркентскую котловину, Балхаш-Алакульскую впадину.
Флора этой провинции относительно молодая и содержит большое число бореальных элементов. Есть эндемичные роды, например зонтичные Pastinacopsis и Seselopsis, но видовой эндемизм не очень высокий. Растительный покров состоит из полынно-злаковых полупустынь, ковыльных степей, лиственничных лесов, елово-пихтовой тайги, высокогорных лугов и кобрезиевых «ковров».
Монгольская провинция охватывает бассейны р. Хобдо и оз. Урэг-Нур, систему хребтов Монгольского Алтая, включая Тайширын-Ула и Адж-Богдо и предгорья Большого Хингана, Среднюю Халху, Восточную Монголию, котловину больших озер, Долину озер, Гобийский Алатау, Восточную, Западную и Алашаньскую Гоби, Ордос, Хэси, Кашгарию, Такла-Макан, Нол-Норскую долину, Цайдам.
Тибетская провинция. Западная граница, имеющая довольно сложную конфигурацию, проходит приблизительно по оси 73° в. д.; северную границу образует Заалай-ский хребет, а южную — Восточный Гунду-куш; далее на восток южная граница проходит по Каракоруму и Гималаям; в восточной части граница идет дугообразно на северо-восток, огибая с юга горы Баян-Хара-Ула. Таким образом, в провинцию входят Памир, Восточный Гиндукуш, Каракорум и большая часть Тибетского нагорья вместе с Цинхаем, занимающим северо-восточную окраину нагорья. На территории СССР эта провинция заходит лишь небольшой частью, занимая Восточный Памир. Флора провинции небогатая и насчитывает, вероятно, не более 1000 видов. Это самая молодая флора во всей Центральноазиатской подобласти, так как ее история начинается, в сущности, лишь после четвертичного оледенения. В этом отношении она напоминает флору Арктики. Основу растительного покрова Тибетской провинции в целом составляют виды центральноазиатского или вообще восточно-древнесредиземноморского происхождения. Виды восточноазиатского происхождения играют значительную роль только в восточной и юго-восточной части Тибетского нагорья.
С. Мадреанское (Сонорское) подцарствоФлора Юго-Западной Северной Америки и Мексиканского нагорья настолько резко отличается от Бореального и Древнесредиземномор-ского подцарств, что вполне может рассматриваться как отдельное, самостоятельное подцар-ство. Флора Мадреанского подцарства развивалась независимо от флоры Древнесредизем-номорского подцарства, но в некоторых отношениях их развитие шло конвергентно. Есть ряд общих родов, например Arbutus, Cercis, Crataegus, Cupressus, Juniperus, Lonicera, Pinus, Pistacia, Platanus, Populus, Prunus, Quercus, Rhamnus, Rhus, Rosa, Rubus, Styrax, Salvia, Viburnum и др., что объясняется скорее всего их происхождением от близких бореальных и отчасти тропических предков. В подцарстве лишь одна область — Мадреанская.
9. Мадреанская (Сонорская) область.
Эта область простирается от Юго-Западного Орегона через Калифорнию до северной части Нижней Калифорнии и включает жаркие пустыни от Южной Калифорнии до Аризоны, Нью-Мексико и Техаса и далее к югу до Мексиканского нагорья, большую часть Невады и Юта и отдельные части соседних штатов.
Флора Мадреанской области характеризуется рядом эндемичных семейств (Crossosomataceae, Koeberliniaceae, Fouquieriaceae, Simondsiaceae и Pterostemonaceae). Это говорит о том, что очень длительное время она развивалась изолированно. Много эндемичных родов (около 10%), в том числе Umbellularia, Anemopsis, Chrysolepis, Romneya, Dendromecon, Carnegiea, Lyonothamnus, Adenostoma, Sarcodes, Allotropa, Dasylirion, Beschorneria.
Видовой эндемизм составляет, вероятно, около 40%.
9. Палеотропическое царство
Палеотропическое царство охватывает тропики Старого Света, за исключением Австралии. В это царство входят все тропические острова Тихого океана, кроме немногих островов, расположенных вдоль побережья Южной Америки.
Богатейшая флора Палеотропического царства включает около сорока эндемичных семейств, из которых мы назовем только пять наиболее известных: непентесовые (Nepenthaceae), диптерокарповые (Dipterocarpaceae), банановые (Musaceae, в узком смысле слова), флагеллариевые (Flagellariaceae) и панданусовые (Panclanaceae). Число родов и особенно видов огромно.
Раскинувшееся на огромное пространство от Африки до Полинезии, Палеотропическое царство очень дифференцировано, и в его пределах можно выделить 5 подцарств: Африканское, Мадагаскарское, Индо-Малезийское, Полинезийское и Новокаледонское.
А.Африканское подцарство
В это подцарство входит большая часть Африканского континента, тропические пустыни Аравийского п-ова, тропические пустыни Ирана, Пакистана и Северо-Западной Индии. Оно может быть разделено на области: Судано-Анголезскую, Гвинео-Конголезскую, область Намиба—Карру и область о-вов Св. Елены и Вознесения.
10. Судано-Анголезская область.
Эта область охватывает тропические части Сахары от Атлантического океана до Красного моря, всю обширную территорию саванн от Сенегала до Судана, Северо-Восточную и Восточную Тропическую Африку, о. Сокотра, тропические части Аравийского п-ова (за исключением юго-западных районов), тропические пустыни Южного Ирана, Пакистана и Северо-Западной Индии, Мозамбик, Южную Родезию, Замбию, Трансвааль, Наталь, части Ботсваны, большую часть Анголы, часть Намибии, Оранжевую провинцию и большую часть Капской провинции. Несмотря на свои колоссальные размеры, Судано-Анголезская область имеет лишь несколько маленьких эндемичных семейств и сравнительно немного эндемичных родов, но очень много эндемичных видов. Особенно характерны виды Acacia (мимозовое) и Commiphora (анакардиевое). Обширные пространства заняты различного типа злаковниками и саваннами, в частности сухой колючекустарниковой саванной. С ними чередуются рощи и массивы смешанных и листопадных лесов и редколесий. Среди деревьев саванны обращают на себя внимание толстоствольные баобабы (Adansonia digitata).Для увлажненных долин характерны галерейные леса. В наиболее сухих районах саванна переходит в тропическую пустыню. В пределах Судано-Анголезской области различаются 4 подобласти.
10а. Сахело-Суданская подобласть
тянется от Сенегала и Гамбии до Судана включительно, занимая части Уганды и Западной Кении. Флористически она бедна, и в ней мало эндемиков. Наиболее характерны виды Acacia и Commiphora, а в менее аридных частях — бобовое Isoberlinia.10b. Нубийско-Раджастанская (Нубийско-Синдская) подобласть.
В эту большую область, расположенную на двух континентах, входят Нубийская пустыня, часть Аравийского п-ова, тропические пустыни Ирана, Пакистана и Северо-Западной Индии. Небольшие эксклавы нубийско-раджастанской флоры имеются в Вади-Эль-Араба в Южной Палестине, в нижней части долины Иордана и на побережье Мертвого моря.
Эта подобласть флористически богаче Сахело-Суданской подобласти, и число эндемиков значительно больше. Из эндемичных или почти эндемичных родов отметим гвоздичное Cometes (от Северо-Восточной Африки и Эфиопии до Северо-Западной Индии), крестоцветные Physorrhynchus (от Южного Ирана до Северо-Западной Индии) и Dipterygium (от Египта до Пакистана), бобовое Taverniera (от Северо-Восточной Африки до Северо-Западной Индии), сапотовое Reptonia (Аравия, Афганистан, Северо-Западная Индия), норичниковое Omania (Аравия), сложноцветные Grantia (Аравия и Иран) и Platychaeta (от Южной Аравии до Северо-Западной Индии), пальма Nannorrhops (от Аравии до Северо-Западной Индии). Характерный для этой подобласти род Ochradenus (резедовое) встречается также на о. Сокотра. Для подобласти характерны также виды родов Acacia, Anogeissus, Balanites, Calligonum, Capparis, Cenchrus, Cleome, Convolvulus, Cordia, Crotalaria, Euphorbia, Fagonia, Jaubertia, Heliotropium, Grewia, Indigofera, Maerua, Leptadenia, Melhania, Pergularia, Tephrosia, Ziziphus, а также Calotropis procera, Gisekia pharnaceoides, Lawsonia inermis, Moringa aptera, Neurada procumbens, Olea cuspidata, Periploca aphylla, Rhazya stricta, Salvadora persica, S. oleoides и др. Многие из этих растений проникают в Сахаро-Ара-вийскую подобласть.
10с. Эфиопско-Йеменская подобласть. Сюда входят Эфиопия, Сомали на юг до Центральной Кении и Танзании, юго-западная часть Аравийского п-ова (Йемен, Асир, НДРИ) и о. Сокотра.
Для подобласти характерны одно эндемичное-семейство (Barbeyaceae) и около 50 эндемичных родов, большей частью олиго- и монотипных, в том числе Sevada, Hagenia, Afrovivella, Саrania, Kissenia, Cephalopentandra, Dendrosicyos, Socotora, Poskea, Drakebrockmania, Wissmannia. Около половины эндемичных родов приурочено к о. Сокотра, флора которого чрезвычайно самобытна и включает много интереснейших растений, в том числе второй вид рода Punica — P. protopunica. Род Goelocarpum (Verbenaceae) является общим для Сокотры (1 вид) и Мадагаскара (4 вида). Для названных районов характерны виды родов Acacia и Commiphoга. Северная часть области включает большую территорию горной флоры, характеризующуюся видами Olea, Ocotea, Hagenia abyssinica и хвойными Juniperus и Podocarpus, а в высокогорьях — древовидными Senecio и Lobelia и кустарниковыми видами Alchemilla и Helichrysum.
10. Анголо-Танзанийская подобласть.
Эта подобласть охватывает Танзанию, Южный Заир, большую часть Анголы, Замбию, Малави, Мозамбик, Южную Родезию, Трансвааль, Наталь, часть Ботсваны, часть Намибии, большую часть территории ЮАР (до Капского царства). Флора характеризуется лишь небольшим числом эндемичных родов. Характерны виды Brachystegia, Julbernardia, Acacia, Commiphora, Terminalia, Combretum и Pleiotaxis, a также Kirkia acuminata, Baikiaea plurijuga и Colophospermum mopane. На юго-востоке значительные площади злаковников с Themeda. В прибрежной полосе Восточной Африки очень узкие и прерывистые пространства характеризуются значительным числом эндемиков, показывающих родовое родство с флорой Мадага-скарской области.
11. Гвинео-Конголезская область.
Гвинео-Конголезская область, или Западно-Африканская область дождевых лесов тянется от Гвинеи до Камеруна, Габона и бассейна реки Конго (Заир).
Флора этой области очень богата. В ней несколько эндемичных семейств (Dioncophylla-ceae, Scytopetalaceae, Octoknemaceae, Medu-sandraceae, Hoplestigmataceae), много эндемичных родов и очень большое число эндемичных видов.
12.Область Намиб — Карру.
Эта сравнительно небольшая область включает западную часть Юго-Западной Африки (включая значительную часть Намибии) и Карру (аридные плато и межгорные равнины, лежащие к югу от р. Оранжевой). В северной части пустыни Намиб и в юго-западной части Анголы есть одно монотипное эндемичное семейство вельвичиевых (Welwitschi-асеае), относящееся к голосеменным растениям и очень изолированное в систематическом отношении. Там же эндемичный род Acanthosicyos — древовидное растение из семейства тыквенных, произрастающее на песчаных дюнах и возникшее в процессе эволюции от лиановых предков. Флора области очень богата эндемичными видами. Особенно характерны виды Mesembryanthemum и близких родов Tetra-gonia, Pelargonium, Rhigozum, а также Pentzia, Pteronia и других кустарниковых сложноцветных.
13. Область островов Св. Елены и Вознесения.
Несмотря на небольшие размеры этих двух вулканических островов, их флора столь своеобразна, что обычно выделяется в самостоятельную область. В то время как о. Вознесения был в момент его открытия почти лишен растительности и в настоящее время имеет лишь два эндемичных вида цветковых растений (Euphorbia origanoides и Hedyotis adscensionis), о. Св. Елены был сплошь покрыт густыми лесами, которые были уничтожены козами и другими домашними животными, завезенными сюда португальцами. Несмотря на то что первоначальная флора была небогата по числу родов и видов, она характеризовалась исключительно высоким видовым эндемизмом. Из 39 видов цветковых растений, сохранившихся на о. Св. Елены, 38 эндемичны, хотя из 28 родов эндемичны только 5. Флора острова обнаруживает наибольшие связи с флорой Африки, особенно Южной Африки, где представлены 22 из 28 родов его цветковой флоры. Что касается эндемичных родов, то они обнаруживают следующие связи: Nesiota (Rhamnaceae) наиболее близка к южноафриканскому роду Phylica, родственному южноамериканским родам; Mellisia (Solanaceae) родственна центрально- и южноамериканскому роду Saracha (от Мексики до Перу); древовидные сложноцветные Commidendron и Melanodendron родственны южноамериканским родам из трибы Asteraae; древовидное сложноцветное Petro-bium, возможно, родственно полинезийским родам Fitchia и Oparanthus.
В.Мадагаскарское подцарство
Сюда входит лишь одна область — Мадагаскарская.
14. Мадагаскарская область.
Область охватывает Мадагаскар, острова Коморские, Альдабра, Амирантские, Сейшельские и Маскаренские. Флора этой области чрезвычайно своеобразна и характеризуется исключительно высоким эндемизмом (85%). В составе мадагаскарской флоры 9 эндемичных семейств (Didymelaceae, Didiereaceae, Diegodendraceae, Asteropeiaceae, Medusagynaceae, Sarcolaenaceae, Rhopalocarpaceae, Melanophyllaceae, Geosiridaceae), большинство которых очень изолированы в систематическом отношении. Имеется около 300 эндемичных родов, большинство из них встречается на самом Мадагаскаре. По числу видов первое место во флоре области занимают орхидные, затем следуют сложноцветные, молочайные, осоковые и злаки.
С. Индо-Малезийское подцарство
Индо-Малезийское подцарство характеризуется исключительно богатой флорой, в составе которой очень много примитивных групп, особенно цветковых растений. По всем данным здесь сосредоточена наиболее древняя на Земле флора цветковых растений.
15. Индийская область.
В Индийскую область входят п-ов Индостан, большая часть Индо-Гангской равнины (за исключением западных частей, относящихся к Судано-Анголезской области), Шри Ланка (Цейлон), Лаккадивские и Мальдивские о-ва и архипелаг Чагос. На севере граница области доходит до субтропической зоны Гималаев.
Эндемичные семейства отсутствуют, и только около 50 эндемичных родов, большинство которых к тому же монотипные и имеют очень локальное распространение. Поэтому флору Индийской области нельзя считать очень самобытной. Но видовой эндемизм очень высокий, особенно на п-ове Индостан, где число эндемичных видов достигает 2100. Естественная растительность Индийской области сильно изменена человеком, а во многих наиболее густонаселенных местах почти не сохранилась. На всем западном побережье п-ова Индостан, от Бомбея до южной его оконечности, в юго-западной части Шри Ланка, в низовьях Ганга и Брахмапутры произрастают тропические дождевые леса, для которых характерны виды диптерокарпуса (Dipterocarpus, семейство диптерокарповых). Сравнительно большие пространства занимают полулистопадные и листопадные сезонные леса (называемые часто муссонными), в частности садовые леса (сал — Shorea robusta, семейство диптерокарповых), сохранившиеся главным образом в виде узкой полосы вдоль южного подножия Гималаев, тиковые леса (тик — Tectona grandis, семейство вербеновых), занимающие большие пространства в Западных Гатах и в центральной части п-ова Индостан, а также смешанные леса. Огромные пространства занимают редколесья, заросли колючих кустарников, саванны и злаковники, на месте которых в прошлом произрастали леса. Вдоль полосы мелководья у морских берегов, защищенных от силь- ного прибоя и периодически затопляемых морем, развивается мангровая растительность. По устьям рек, за исключением рек, впадающих в Аравийское море, по лагунам и прибрежным болотам с внутренней стороны мангровых развиты заросли бесстебельной пальмы нипа (Nypa fruticans).
16. Индокитайская область.
В эту область входят большая часть Ассама, п-ов Индокитай, Андаманские и Никобарские о-ва, тропические районы Южного Китая, о. Хайнань. Эндемичные семейства отсутствуют, но более 250 эндемичных родов.
Естественная растительность Индокитайской области сохранилась лучше и значительно богаче, чем растительность Индийской области. Довольно большие площади занимают тропические дождевые леса, в которых очень заметную роль играют различные представители семейства диптерокарповых. В горах тропический дождевой лес постепенно переходит в леса субтропического типа (так называемые мшистые леса), для которых характерны вечнозеленые представители семейства дубовых и лавровых, крупные древовидные рододендроны и другие вересковые, магнолиевые, чайные и др. На стволах и ветвях деревьев мшистого леса растут многочисленные эпифиты (папоротники, орхидные и др.); особенно большую роль среди эпифитов играют мхи и печеночники, густо покрывающие ветви деревьев. Местами, особенно выше в горах, значительную роль играют хвойные, в том числе подокарпусы. В районах с ясно выраженным засушливым сезоном распространены полулистопадные и листопадные сезонные леса, занимающие очень большие площади в Центральной Бирме, Таиланде и в сухих внутренних районах Лаоса. Доминирующей породой в этих лесах нередко является тик (особенно в Северной Бирме и в Северном Таиланде) или же низкорослые и листопадные виды диптерокарповых. В относительно сухих местообитаниях, начиная с уровня моря, встречаются сосновые леса. На месте уничтоженных сезонных лесов появляются вторичные саванны, редколесья, кустарники и заросли бамбука. На морских побережьях и в дельтах местами распространена мангровая растительность и заросли нипы.
17. Малезийская (Малайская) область.
Сюда входят п-ов Малакка, острова Ява, Суматра, Калимантан, Филиппины. Флора характеризуется двумя эндемичными семействами (Tetrameristaceae и Scyphostegiaсеае) и многими эндемичными родами. Для растительности Малезийской области наиболее характерны роскошные тропические дождевые леса, в которых большую роль играют диптерокарповые, гигантские фикусы, пальмы, панданусы, многочисленные папоротники. В горах развиты мшистые леса, которые выше переходят в криволесье и заросли кустарников. Имеются также сезонные леса, редколесья и вторичные саванны. Характерна мангровая растительность.
18. Папуасская область.
В эту область входят острова Сулавеси, Тахуна и Талауд, Молуккские, Банда, Танимбар(?), Ару, Мисоол, Сала-вати, Вайгео, Новая Гвинея, Адмиралтейства, арх. Бисмарка, о. Новая Британия, о-ва Тро-бриан, Муруа, Д'Антркасто, арх. Луизиада, Соломоновы о-ва. Эндемичные семейства отсутствуют, число эндемичных родов невелико, но видовой эндемизм высокий.
<Н3>19. Фиджийская область.Область охватывает острова Санта-Крус, Новые Гебриды, Фиджи, Тонга, Самоа. Во флору области входят примитивное эндемичное семейство Degeneriaceae, около 15 эндемичных родов и более 70% эндемичных видов.
D. Полинезийское царство
В Полинезийское подцарство входят области с чисто островными флорами, лишенными эндемичных семейств, но характеризующиеся часто высоким родовым и видовым эндемизмом. Исторически полинезийская флора является производной от Индо-Малезийской.
20. Гавайская область.
Эта небольшая область охватывает Гавайские о-ва и атолл Джонстона.
Флора Гавайской области содержит около 15% эндемичных родов сосудистых растений, в том числе гвоздичное Schiedea, фиалковое Isodendrion, бегониевое Hillebrandia, губоцветное Stenogyne, мареновое ВоЬеа, колокольчиковые Clermontia, Cyanea, Delissea, Rollandia и Trematolobelia, сложноцветное Hesperomannia. Особенно характерны древовидные роды из семейства колокольчиковых. Более 90% видов коренной гавайской флоры эндемичные.
21. Полинезийская область.
Полинезийская область охватывает острова Каролинские, Марианские, Маркус, атолл Уэйк, Маршалловы, Науру, Ошен, Гилберта, Хауленд, Бейкер, Эллис, Феникс, Увеа (Уоллис), Хорн, Токелау, Лайн (Центральные Полинезийские Спорады), Кука (северные), Общества, Тубуаи, Рапа, Туа-моту, Маркизские, Мангарева (Гамбье), Питкэрн, Хендерсон, Дюси, Пасхи, Сала-и-Гомес. К числу немногочисленных эндемичных родов относятся аралиевое Reynoldsia, тутовое Metatrophis, липовое Tahitia (о-ва Общества), геснериевое Cyrtandroidea (Маркизские о-ва), мареновые Hitoa (Таити) и Sarcopygme (Самоа), колокольчиковые Sclerotheca (о-ва Кука, Общества) и Apetahia (о-ва Общества, Маркизские, Рапа), сложноцветное Fitchia и пальма Solfia (Самоа).
Е. Новокаледонское подцарство
Новокаледонское подцарство отличается столь большим своеобразием флоры, что, безусловно, заслуживает выделения в отдельное под-царство. Оно состоит лишь из одной области.22. Новокаледонская область.
В Новокаледонскую область входят острова Новая Каледония, Пен (Куние) и Луайоте.
Область характеризуется несколькими эндемичными семействами (Amborellaceae, Strasburgeriaceae, Oncothecaceae, Paracryphiaceae и Phellineaceae) и около 110 эндемичными родами, в том числе чрезвычайно своеобразными папоротником из семейства глейхениевых (Stromatopteris), хвойными Austrotaxus (иногда выделяется в отдельное семейство), Parasitaxus (единственное паразитное растение среди хвойных), Prumnopitys и Neocallitropsis, цветковыми Exospermum (примитивный род из бессосудистого семейства винтеровых), Canacomyria (восковниковое), Осеаnоpapaver (каперсовое), Maxwellia (баобабовое), Myodocarpus (аралиевое с плодами, как у зонтичных). Видовой эндемизм достигает 85%. Нет на Земле другой территории такого же размера, которая могла бы сравниться с Новой Каледонией столь большим числом эндемичных семейств и родов. По проценту эндемичных родов с Новой Каледонией можно сравнить лишь о-ва Гавайские и Хуан-Фернандес. Интересно, что далеко не все эндемичные роды монотипны и число видов некоторых из них достигает нескольких десятков. Это указывает на то, что здесь длительное время шли интенсивные процессы видообразования. В то же время во флоре Новой Каледонии имеется значительное число древних и примитивных родов, и здесь встречается 6 из 11—12 известных бессосудистых родов цветковых растений (Amborella, Belliolum, Bubbia, Drimys, Exospermum и Zygogynum), причем три из них (Amborella, Exospermum и Zygogynum) эндемичны. По родовому составу флоры Новая Каледония имеет много общего с флорой островов Новая Гвинея, Соломоновых, Новых Гебрид и Фиджи, с одной стороны, и с флорой Северо-Восточной Австралии — с другой.
III. Неотропическое царствоНеотропическое царство занимает южные тропические части п-овов Калифорния и Флорида, низменности и побережья Мексики, всю Центральную Америку с Антильскими о-вами, большую часть Южной Америки (за исключением южных умеренных и субтропических ее частей, относящихся к Голантарктическому царству) и ряд тропических островов, примыкающих к Южно-Американскому континенту. Таким образом, это флористическое царство полностью располагается в Новом Свете, почему и называется неотропическим.
Неотропическая флора имеет общее происхождение с палеотропической, и можно предполагать, по крайней мере в отношении цветковых растений, что она уходит корнями в Па-леотропическое царство. Имеется много общих семейств и даже родов с так называемым пан-тропическим распространением, т. е. растений, распространенных в тропиках как Старого, так и Нового Света (иногда выходя за пределы тропиков). Назовем, например, семейства аноновых (Annonaceae), гернандиевых (Hernandiaсеае), лавровых (Lauraceae), перцевых (Pipeгасеае), крапивных (Urticaceae), диллениевых (Dilleniaceae), страстоцветных (Passifloraceae), баобабовых (Bombacaceae), молочайных (Еnphorbiaceae), ризофоровых (Rhizophoraceae), миртовых (Myrtaceae), анакардиевых (Апасаrdiaceae), сапиндовых (Sapindaceae), мальпигиевых (Malpighiaceae), протейных (Proteaceae), бигнониевых (Bignoniaceae), орхидных (Orchidaceae), пальм (Arecaceae). Довольно много также общих родов (вероятно, не менее 450).
Все это говорит о том, что в течение значительного времени была тесная миграционная связь между тропиками Старого и Нового Света; при этом миграция происходила не только из Старого Света в Новый, но и частично в противоположном направлении.
Есть основания предполагать, что некоторая связь Южной Америки с тропической Западной Африкой продолжалась вплоть до середины верхнего мела. Одним из наиболее замечательных примеров былой связи флоры тропической Западной Африки с флорой тропической Южной Америки является распространение семейства бромелиевых (Bromeliaceae). Почти все виды семейства бромелиевых произрастают в тропической Южной Америке, за исключением только Pitcairnia feliciana, эндемичного для Гвинеи.
Совершенно аналогично распространение другого неотропического семейства — Rapateaceae, большинство родов которого произрастает в тропической Южной Америке, за исключением рода Maschalocephalus, эндемичного для Либерии. Однако отделение Южной Америки от Африки произошло достаточно давно, и поэтому неотропическая флора в течение очень длительного времени развивалась самостоятельно и выработала около тридцати эндемичных семейств и огромное множество эндемичных родов и видов. Неотропическое царство подразделяется на несколько областей.
23. Карибская область.
Сюда входят южная тропическая часть п-ова Калифорния, низменности и побережья Мексики, самая южная тропическая часть п-ова Флорида, острова Ревилья-Хихедо, Клиппертон, Флорида-Кис, Багамские, Бермудские, Большие и Малые Антильские, Гватемала, Панама, Северная Колумбия, Северная Венесуэла и о. Тринидад. Во флору области входят 2 эндемичных семейства (Picrodendraceae и Plocospermataceae) и более 500 эндемичных родов (на Бермудских о-вах эндемичные роды отсутствуют). Очень высокий процент эндемичных видов.
24. Область Гвианского нагорья.
Область Гвианского нагорья характеризуется древней и оригинальной флорой, насчитывающей около 100 эндемичных родов, процент эндемичных видов достигает 70, а в высокогорьях — даже от 90 до 95. Во флоре Гвианского нагорья представлен ряд примитивных семейств, в том числе магнолиевые (несколько видов рода Magnolia) и винтеровые (Drimys brasiliensis).
25. Амазонская область.
Амазонская область включает низменности бассейна Амазонки, побережье Бразилии и Гвианы, острова Фер-нанду-ди-Норонья и Сан-Паулу.
Богатейшая флора Амазонской области содержит лишь одно эндемичное семейство Dialypetalanthaceae, но в ней имеется не менее 500 эндемичных родов и не менее 3000 эндемичных видов. Наиболее характерной чертой растительности Амазонской области является роскошный дождевой тропический лес. Нигде в мире дождевые тропические леса не занимают столь большой площади и не отличаются таким богатством растительных форм, как в бассейне Амазонки. Из наиболее известных полезных растений этой области, широко культивируемых в тропических странах, нужно указать каучуконосное дерево гевею (Hevea brasiliensis) и шоколадное дерево (Theobroma cacao).
26. Центральнобразильская область.
Область занимает Бразильское нагорье, Гран-Чако и острова Триндади и Мартин-Вас. Эндемичные семейства отсутствуют, но имеются около 400 эндемичных родов, в том числе Diclidanthera из семейства истодовых.
27. Андийская область.
Вытянутая с юга на север, область охватывает западные береговые хребты и побережье Южной Америки от Колумбии до пустыни Атакама, Галапагосские о-ва и о. Кокос.
Область характеризуется двумя эндемичными семействами: Malesherbiaceae (также в Аргентине) и Nolanaceae (на севере Чили и в Перу). Во флору области входит большое число эндемичных родов. Из наиболее известных андий- ских, эндемичных родов упомянем хинное дерево (Cinchona). Андийская область является родиной многих важных культурных растений.
IV. Капское царство
Капское царство — самое маленькое среди флористических царств Земли, но благодаря исключительному своеобразию флоры и самостоятельности ее развития все географы растений единодушно отделяют его от остальной флоры Африки. Оно состоит лишь из одной области — Капской.
28. Капская область.
В Капскую область входит южная оконечность Африки от Кланвильяма на западе до окрестностей Порт-Элизабет на востоке. Несмотря на небольшие размеры области, ее флора необыкновенно богата (около 7000 видов) и насчитывает несколько эндемичных семейств (Grubbiaceae, Roridulaceae, Bruniaceae, Penaeaceae, Greyiaceae, Geissolomataceae, Retziaceae). Более 280 родов имеют своим центром Капскую область, а более 210 из них эндемичны для области (включая также роды эндемичных семейств).
Видовой эндемизм очень высокий, около 90%. Очень характерны многочисленные виды вереска (Erica), солнечника (Mesembryanthemum) и многочисленные протейные (Proteaceae), представленные видами рода Protea и целым рядом эндемичных родов, в том числе Leucadendron. Один из видов этого рода — серебряное дерево (L. argenteum) со сверкающими, блестящими, серебристыми листьями — является одним из наиболее характерных растений капской флоры. Многими видами представлен здесь пеларгониум (Pelargonium, гераниевое), сложноцветные Helichrysum и Senecio и др. Одной из характерных особенностей растений капской флоры является обилие листовых и стеблевых суккулентов и огромное количество геофитных однодольных.
Южная часть Африки отделилась от Южной Америки в раннем мелу, а от Антарктики — в начале третичного периода. Трудно предположить, что до расхождения этих материков уже существовали все те роды цветковых растений, которые являются общими между Капским и Голантарктическим царствами. Более вероятно, что их расселение происходило через соединявшие их архипелаги, т. е. с помощью птиц, ветра и морских течений. Так, общие роды Асаепа (розоцветное), Gunnera (гуннеровое), Bulbinella (лилейное), Leptocarpus (рестионо-вое), очевидно, распространились уже после расхождения южных материков. В то же время таксоны более высокого ранга, такие, как семейства Restionaceae и особенно Proteaceae, вполне могли существовать еще до этого события. В прошлом капская флора занимала в Африке гораздо большую территорию, чем в настоящее время, и из-за возрастающей сухости климата продолжает неуклонно сокращаться. Капская флора не дала человечеству ни одного важного в хозяйственном отношении культурного растения, но она продолжает служить неисчерпаемым источником красивых садовых и комнатных растений: агапантус, амариллис, декоративная спаржа, фризия, гальтония, гер-бера, виды гладиолуса, кливия, виды книпхо-фии, плюмбаго и др.
V. Австралийской царство
Флора Австралии очень самобытна и отличается высоким эндемизмом. В ее флоре имеется целый ряд эндемичных семейств: Austrobaileyaceae, Idiospermaceae, Byblidaceae, Cephalotaceae, Eremosynaceae, Akaniaceae, Tremandraceae, Brunoniaceae и др. Кроме того, Австралийское царство является центром развития таких семейств, как Pittosporaceae, Epac-ridaceae, Stackhoussiaceae, Myoporaceae и Goodeniaceae, а также многих родов. Наибольшую роль во флоре Австралии играют злаки, бобовые, сложноцветные, орхидные, лилейные, молочайные, осоковые, рутовые, миртовые (особенно подсемейство Leptospermoideae) и протейные. В то же время отсутствуют многие широко распространенные группы, например хвощи, подсемейство Maloideae семейства Rosaceae, бамбуки и такие семейства, как чайные (Theaсеае), вересковые (Ericaceae), бегониевые (Веgoniaceae) и др.
В состав флоры входят около 570 эндемичных родов.
Важную роль в растительном покрове Австралии играют многочисленные виды акации (Acacia) и особенно эвкалиптов (Eucalyptus), а также казуарины (Casuarina), некоторые другие миртовые (Melaleuca, Leptospermum и дp.) многочисленные протейные (Proteaceae), особенно виды Banksia. В то время как одни элементы австралийской, флоры, например миртовые, роды Acacia и Casuarina, многочисленные стеркулиевые (Sterculiaceae) и рутовые (Rutaceae), обнаруживают преимущественно палеотропические связи, другие элементы, например протейные (Proteaceae), эпакридовые (Epacridaceae — семейство, как бы заменяющее в Австралии близкое семейство вересковых) или однодольное семейство Restionaceae, обнаруживают широкие связи с флорами южного полушария.
Как предполагают, движение Австралии к северу началось в середине мелового периода и в эоцене (45—49 млн. лет назад), окончательно прервалась ее связь с Южной Америкой через Антарктику. Австралия по мере движения к северу постепенно вошла в зону более теплого климата. Флористический обмен между Австралией и Антарктикой, а через Антарктику и с южными областями Южной Америки продолжался вплоть до миоцена, чем и объясняется наличие многих общих таксонов не только араукариевых, подокарповых, винтеровых, протейных, рестионовых и других характерных семейств южного полушария, но и многих родов, например Nothofagus, Hebe, Donatia, Drapetes, причем многие из них являются общими с новозеландской флорой. В большинстве случаев эти общие элементы представляют собой реликты древней голантарктической флоры, которые в пределах Австралийского царства лучше всего сохранились в горах Юго-Восточной Австралии и в Тасмании. В то же время на родовом уровне очень мало общего с капской флорой. Первоначальное ядро флоры Австралийского царства возникло в результате преобразования элементов древней голантарктической флоры, эволюция которых в результате длительной изоляции Австралии и сложившихся в ней своеобразных физико-географических условий шла в основном в сторону ксерофилизации. Ксеро-фильная флора Австралии формировалась по мере вступления Австралии в зону низких осадков южной окраины тропиков, т. е. в основном после эоцена, хотя семиаридные условия, по некоторым данным, существовали еще в позднем мелу. По мере движения Австралии к северу она все больше входила в сферу влияния палеотропиче-ской флоры, некоторые представители которой дают начало таким характерным элементам австралийской флоры, как акации и эвкалипты.
В Австралийском царстве различаются три ясно выраженные флористические области.
29. Северо-Восточно-Австралийская область.
Область охватывает северные, восточные и юго-восточные лесные и отчасти саванновые районы Австралии вместе с прибрежными островами и о. Тасмания. Во флору области входят 5 эндемичных семейств (Austrobaileyaceae, Tetracarpaeaceae, Petermanniaceae, Idiospermaceae и Akaniaceae) и более 150 эндемичных родов. Тасмания имеет 14 эндемичных родов, в том числе хвойные Athrotaxis, Diselma и Microcachrys и цветковые Tetracarpaea, Prionotes, Isophysis.
30. Юго-Западно-Австралийская область.
Во флору области входят 3 эндемичных семейства (Cephalotaceae, Eremosynaceae и Emblingiaсеае) и около 125 эндемичных родов (включая Dryandra, Nuytsia, Stirlingia и др.). Очень высок видовой эндемизм (75% или больше).
31. Центральноавстралийская, или Эремей-ская, область. Область охватывает северные и восточные саванновые районы, центральные пустыни и Южную Австралию.
Во флоре области эндемичные семейства отсутствуют, но имеется около 40 эндемичных родов, многие из которых относятся к семействам маревых, крестоцветных и сложноцветных.
VI. Голантарктическое царство
Голантарктическое флористическое царство относительно самое бедное в ряду флористических царств Земли и по богатству своей флоры значительно уступает Голарктическому царству. Тем не менее оно содержит одиннадцать небольших моно- или олиготипных эндемичных семейств (Lactoridaceae, Gomortegaceae, Hectorellaceae, Halophytaceae, Francoaceae, Aextoxicaceae, Tribelaceae, Griseliniaceae, Misodendraceae, Alseuosmiaceae и Donatiaceae) и значительное число эндемичных родов, из которых многие характеризуются очень разорванным ареалом. Несмотря на то, что территории, входящие в состав Голантарктического царства, значительно удалены друг от друга (например, о-ва Хуан-Фернандес и Огненная Земля отделены от Новой Зеландии и о. Лорд-Хау колоссальными морскими пространствами), в составе их флоры имеется много общих родов и близких и даже идентичных видов. Наличие многих общих таксонов между о-вами Хуан-Фернандес и умеренной Южной Америкой, антарктическими островами и Новой Зеландией с прилегающими островами, а также Тасманией и горами Юго-Восточной Австралии и отчасти Южной Африки уже давно привело к выводу о существовании в прошлом древнего центра умеренной флоры в южном полушарии. Еще в 1853 г. Дж. Хукер отметил ботаническое родство между внетропической Южной Америкой, антарктическими островами, Новой Зеландией и Тасманией. В настоящее время существование древней умеренной голантарктической флоры, некогда единой, но впоследствии разорванной, не вызывает сомнений.
Расцвет голантарктической флоры восходит к тем временам, когда ныне разрозненные части территории Голантарктического царства были соединены в единую сушу, составляющую часть южного материка Гондваны. Разделение этой суши на отдельные, постепенно удалявшиеся друг от друга части произошло главным образом в течение первой половины третичного периода, когда цветковые растения были уже господствующей группой в растительном мире Земли. Во время плейстоценовых оледенений многие элементы голантарктической флоры продвинулись далеко на север, особенно в Южной Америке, где в высокогорной флоре Анд они достигли Эквадора и Колумбии. Сюда относятся, например, роды Colobanthus (гвоздичное), Асаепа (розоцветное), Azorella (зонтичное) и Ourisia (норичниковое). В это время целый ряд голарктических растений смог благодаря снижению снеговой линии пройти по низким горам Панамского перешейка и дойти до Южного Чили. Но голарктический элемент проник в Голантарктику еще в третичное время, что доказывается наличием в субантарктической Южной Америке и Новой Зеландии хорошо обособленных эндемичных видов и даже надвидовых таксонов (вплоть до секций) таких родов, как Caltha, Ranunculus, Berberis, Stellaria, Rumex, Draba, Geum, Hydrangea, Saxifraga, Veronica, Euphrasia, Pedicularis, Plantago, Juncus, Luzula, Poa и др.
Еще в начале мелового периода Антарктика, Южная Америка (вместе с Фолклендским плато), Африка и Мадагаскар, Индия, Австралийская плита (Австралия, Новая Гвинея, погруженное Квинслендское плато и Тасмания), Новая Каледония и Новая Зеландия вместе с почти полностью погруженным плато Кэмпбелла и островами и погруженными хребтами Лорд-Хау, Норфолк и Маккуори составляли гигантский южный континент Гондвану, отделенный морем от северного континента Лавразии. В южной умеренной и субтропической зонах Гондваны формировалась голантарктическая флора, которая развивалась параллельно северной голарктической флоре. Но со временем Гондвана стала распадаться на отдельные части, которые медленно раздвигались в разные стороны.
Как геологические, так и биогеографические данные указывают на то, что от Гондваны очень давно начала отделяться Африка (вместе с Мадагаскаром), которая в конце мелового периода уже находилась на значительном расстоянии от Южной Америки и Антарктики. Поэтому голантарктический элемент представлен в Южной Африке, на Мадагаскаре и о. Реюньон слабее, чем в других умеренных и субтропических странах южного полушария. Тем не менее имеется немало общих таксонов между Африкой — Мадагаскаром — Реюньоном и другими частями бывшей Гондваны. Одним из наиболее замечательных примеров служит род Dietes (ирисовое), 3 вида которого произрастают в Южной Африке, а один (D. robinsoniana) — на о. Лорд-Хау.
В позднем мелу (около 80 млн. лет назад) от Восточной Антарктики, представлявшей собой архипелаг, отделилось плато Кэмпбелла (включая Новую Зеландию), а в эоцене (45— 50 млн. лет назад) стала отходить в северном направлении Австралия, и тем самым прервалась ее связь с Южной Америкой через Антарктику. В то время как Австралия передвинулась на север на 15° от Индийско-Антарктического поднятия, Антарктика продвинулась на 15° к югу. Последующее развитие биоты (флоры и фауны) Австралии пошло самостоятельными путями, и голантарктический элемент сохранился главным образом в горах Юго-Восточной Австралии и Тасмании, в то время как в Новой Зеландии и на окружающих ее островах он продолжал преобладать.
Антарктика приняла положение, близкое к современному, и ее растительный мир сильно обеднел. В Голантарктическое царство входят 4 области: Хуан-Фернандесская, Патагонская, область субантарктических островов и Новозеландская.
32. Хуан - Фернандесская область.
Область охватывает о-ва Хуан-Фернандес (Александр-Селькирк, Робинзон Крузо и Санта-Клара) и Десвентурадас (Сан-Феликс и Сан-Амбросио). Острова представляют собой вулканические пики на подводном хребте Хуан-Фернандес.
Во флоре области одно уже почти вымирающее эндемичное семейство Lactoridaceae и 20 эндемичных родов, в том числе папоротник Thyrsopteris (монотипный род из семейства циатейных), Nothomyrcia (монотипный род из семейства миртовых), Selkirkia (монотипный род из семейства бурачниковых), Cuminia (монотипный род из семейства губоцветных), представители трибы Senecioneae семейства сложноцветных (роды Robinsonia, Rhetinodendron и Symphyochaeta), древовидные васильки (Centaurodendron и Yunquea — монотипные роды, вероятно, производные от Centaurea), целый ряд древовидных представителей трибы Cichorieae семейства сложноцветных, пальма Juania (родственное североандийскому роду Ceroxylon и мадагаскаро-коморскому роду Ravenea), бромелиевые Ochagavia и Hesperogreigia, злаки Podophoms, Pantathera и Megalachne. Около 70% флоры сосудистых растений, насчитывающих около 200 видов, являются эндемичными.
Флористически о-ва Хаун-Фернандес наиболее близки к Патагонской области и поэтому должны быть включены в Голантарктическое царство, а не в Неотропическое, как это принято почти во всех флористических системах. Оледенения повлияли на флору островов Хуан-Фернандес и Десвентурадас гораздо меньше, чем на флору Чили, и поэтому эти острова можно рассматривать как убежище более древней, доплейстоценовой флоры. Обе группы островов заметно отличаются друг от друга флористически. Это объясняется тем, что наиболее богатая лесная флора сохранилась на о. Робинзон-Крузо (Мас-а-Тьерра), а на о. Александр-Селькирк развита альпийская флора. Во флоре Хуан-Фернандесской области имеется ряд изолированных таксонов, таких, как Thyrsopteris elegans, Lactoris fer-nandeziana, некоторые древовидные роды зонтичных и сложноцветных и ряд родов и видов, связанных с родами и видами Новозеландской области, Океании и Гавайских островов.
Есть небольшая группа растений, связанная с андийской флорой: эндемичные роды Selkirkia, Cumina и Juania, эндемичные виды Веrberis и эндемичные древовидные виды Nicotiana.
Наиболее значительным элементом Хуан-Фернандесской флоры является чилийский, хотя эти растения большей частью эндемичны и, как правило, очень своеобразны. Наконец, имеется значительный магелланский элемент (Александр-Селькирк), но большинство видов растений неэндемичны.
33. Патагонская область.
Область охватывает внетропические части Южной Америки от субтропических «монта» Аргентины и араукариевых лесов Южной Бразилии до Огненной Земли, а также о-ва Фолклендские, Эстадос, Диего-Рамирес, Южная Георгия, Южные Шетландские и Южные Оркнейские, Аделейд и Антарктический п-ов. Остров Нени у западного побережья Антарктического п-ова является крайним южным пределом распространения сосудистых растений.
Патагонская область характеризуется довольно разнообразной флорой, состоящей в основном из дериватов древнего голантарктического элемента, но со значительной примесью таксонов голарктического происхождения, а на севере — также неотропических элементов.
Во флоре области 7 эндемичных семейств: Gomortegaceae, Halophytaceae (субаридные районы Аргентины, от Ла-Риоха до Санта-Крус), Malesherbiaceae, Tribelaceae (Южное Чили и Огненная Земля), Francoaceae (Чили), Aexto-xicaceae (Чили) и Misodendraceae (в лесных районах от 33° ю. ш. до Магелланова пролива) и множество эндемичных или почти эндемичных родов, большинство которых приурочено к Чили. Из числа наиболее интересных эндемичных родов отметим папоротник Leptocionium (Hymenophyllaceae), хвойные Saxegothaea (Podocarpaceae), Austrocedrus, Pilgerodendron и Fitzroya (Cupressaceae), цветковые Peumus (Monimiaceae), Boquila и Lardizabala (Lardizabalaceae), Philippiella (Caryophyllaceae), Austrocactus и Holmbergia (Cactaceae), Berberidopsis (Flacourtiaceae), Niederleinia (Frankeniaceae), Lebetanthus (Epacridaceae), Ovidia (Thymelaeaceae), Quillaja и Kageneckia (Rosaceae), Saxifragella (Saxifragaceae), Zuccagnia (Fabaceae), Tepualia (Myrtaceae), Magallana (Tropaeolaceae), Gymnophyton, Laretia и Mulinum (Apiaceae), Talguenea (Rhamnaceae), Schizanthus (Solanaceae), Melosperma и Monttea (Scrophulariacene), Hygea, Mitraria и Sarmienta (Gesneriaceae), Chiliotrichum, Melalema и Nassauvia (Asteraceae), Tetroncium (Juncaginaceae), Gilliesia, Leontochir, Leucocryne, Schickendantziella и Solaria (Liliaceae), Lapageria (Philesiaceaceae), Conanthera и Tecophilaea (Tecophilaeaceae), Tapeinia (Iridaceae), Fascicularia (Bromeliaceae), Ortachne (Poaceae), Jubaea (Arecaceae). Очень много эндемичных видов. Подавляющее большинство эндемиков сосредоточено в Чили, где сохранилось наибольшее число древних голантарктических элементов.
34. Область субантарктических океанических островов.
Эта переходная область между Новозеландской и Патагонской включает о-ва Тристан-да-Кунья вместе с о. Гоф, о-ва ПринсЭдуард, Крозе, Амстердам, Сен-Поль, Кергелен и Хёрд вместе с расположенным близко от него о. Макдональд.
Флора этой области очень обеднена, но, как показывают данные палеоботаники, в прошлом она была значительно богаче, и хвойные и но-тофагусовые леса покрывали обширные пространства. Для области характерны два эндемичных монотипных рода: Pringlea (P. anti-scorbutica) из семейства крестоцветных (острова Принс-Эдуард, Крозе, Кергелен и Хёрд) и Lyallia (L. kergelensis) из очень близкого к гвоздичным семейства Hectorellaceae (Кергелен). Первый род относится к числу примитивных родов семейства крестоцветных и наиболее близок к родам Stanleya (запад США) и Warea (юго-восток США), имеющих много общего с Cleome и близкими родами из семейства каперсовых, а второй — к монотипному новозеландскому роду Hectorella.
Относительно более богата флора о-вов Тристан-да-Кунья, насчитывающая 44 вида сосудистых растений, из которых около 30 эндемичны. Несмотря на большую близость к Южной Африке, чем к Южной Америке, флора области с Южной Америкой связана гораздо теснее. Например, Chevreullia stolonifera (сложноцветное) является общей с ней, а злак Роа flabellata на о. Гоф является магелланским элементом.
На о-вах Сен-Поль и Амстердам насчитывается 17 видов, из которых 7 эндемичны; на о. Принс-Эдуард произрастают 19 видов растений, из которых 2 эндемичны. На о-вах Крозе насчитывается 24 вида, в том числе отсутствующие на о-вах Принс-Эдуард Colobanthus kerguelensis, Montia fontana, Limosella lineate, GaHum antarcticum, Juncus pusillus, Deschampsia antarctica, причем здесь нет ни одного эндемичного вида. Флора о. Кергелен насчитывает 30 видов. По сравнению с флорой о-вов Крозе здесь прибавляются Polypodium vulgare var. eatonii, Ranunculus moseleyi (эндемик), Lyallia kerguelensis (эндемик), Uncinia compacta, Poa kerguelensis и Festuca erecta.
Несмотря на свое географическое положение, флора Кергелена обнаруживает наибольшую близость к флоре Огненной Земли, с которой имеет 17 общих видов. Наконец, на о. Хёрд произрастает всего лишь 8 видов растений: Pringlea anti-scorbutica, Colobanthus kerguelensis, Acaena ad-scendens, Callitriche antarctica, Azorella selago, Deschampsia antarctica, Poa cookii и Р. kerguelensis. Эндемичные виды отсутствуют. Единственный вид, который встречается на всех островах. — Callitriche antarctica.
35. Новозеландская область.
В область входят острова Новая Зеландия, Стыоарт, Те-Снэрс, Три-Кингс, Лорд-Хау и Болс-Пирамид, Норфолк, Кермадек, Чатем, Антиподов, Кэмп-белл, Окленд и Маккуори.
Эндемичные семейства отсутствуют, но имеется 50 эндемичных родов: Loxsoma (Loxsomaceae), Pseudowintera (Winteraceae), Hectorella (Hectorellaceae), Entelea (Tiliaceae), Hoheria (Malvaceae), Corokia (Escalloniaceae), Ixerba (Brexiaceae), Alseuosmia (Alseusomiaceae), Carmichaelia (Fabaceae, Новая Зеландия и Лорд-Хау), Lophomyrtus и Neomyrtus (Myrtaceae), Plectоmirtha (Anacardiaceae, Три-Кингс), Stilbocarpa (Araliaceae, субантарктические острова), Kirkophytum (Araliaceae, Те-Снэрс, Стыоарт и прилегающие островки), Coxella (Apiaceae, Чатем), Lignocarpa и Scandia (Apiaceae), Dactylanthus (Balanophoraceae), Myosotidium (Boraginaceae, Чатем), Parahebe (Scrophulariaceae, род, близкий к Veronica), Negria (Gesneriaceae, Лорд-Хау), Rhabdothamnus (Gesneriaceae), Teucridium (Verbenaceae), Oreostylidium (Stylidiaceae), Pachystegia (Asteraceae, род, близкий к Olearia), Haastia (Asteraceae), Leucogenes (Asteraceae, род, близкий к Helichrysum), Phormium (Liliaceae, Новая Зеландия и Норфолк), Rhopalostylis (Arecaceae, Новая Зеландия, Норфолк и Кермадек), Lepidorrhachis, Hedyscepe и Howeia (Arecaceae, Лорд-Хау), Sporadanthus (Restionuceae, Новая Зеландия и Чатем), Aporostylis (Orchidaceae), Desmoschoenus (Gyperaceae). Видовой эндемизм флоры Новозеландской области очень высокий. У папоротников эндемизм составляет около 40%, у родов Dicksonia и Cyathea он составляет около 75%. Интересно отметить, что папоротниковая флора Новой Зеландии обнаруживает много общего с умеренной Австралией и Тасманией (около 45%), в то же время имеет удивительно мало общего с флорой антарктической Южной Америки. Из немногих общих элементов укажем Sphaerocionium (Hymenophyllum) ferrugineum (наиболее темные части дождевых лесов Новой Зеландии, Чили и Хуан-Фернандес), Grammi-tis billardieri (Новая Зеландия, особенно о. Южный, и Восточная Австралия, Южное Чили, Фолклендские о-ва и Кергелен), Blechnum penna-marina (Новая Зеландия, особенно Южные Альпы, Восточная Австралия, Южное Чили, Тристан-да-Кунья, Кергелен, Крозе и Марион, а также Сен-Поль и Амстердам к северу от Кергелена) и Polystichum mohrioides (Окленд и Кэмпбелл, Фолклендские, Южная Георгия, Марион, Амстердам и Южное Чили). Исключительно высок видовой эндемизм во флоре хвойных, достигающий 100%. Из видовых эндемиков отметим Araucaria heterophylla (Норфолк и Филип), Agathis australis (низменные леса о. Северного, почти от мыса Северного до широты 38°), 7 видов Podocarpus, 6 видов Dacrydium, 3 вида Phyllocladus и 2 вида Libocedrus. Большая часть этих видов приурочена к Новой Зеландии, но некоторые (большая часть видов Podocarpus и Dacrydium) произрастают также на о. Стьюарт. Некоторые из новозеландских видов Podocaprus и Dacrydium имеют викарные виды в Чили. Флора цветковых растений Новозеландской области отличается также высоким видовым эндемизмом (по-видимому, не менее 80%). Наиболее богаты эндемичными видами роды Ranunculus и Clematis (Ranunculaceae), Colobanthus (Caryophyllaceae), Gaultheria (Ericaceae), Dra-cophyllum (Epacridaceae), Myrsine (Myrsina-ceae), Pimelea (Thymelaeaceae), Pittosporum (Pittosporaceae), Alseuosmia (Alseuosmiaceae, эндемичный род), Acaena (Rosaceae), Carmichaelia (Fabaceae, эндемичный род), Metrosideros (Myrtaceae), Epilobium (Onagraceae), Schizellema, Aciphylla и Anisotome (Apiaceae), Gentiana (Gentianaceae), Coprosma (Rubiaceae), Myosotis (Boraginaceae), Euphrasia, Ourisia, Parahebe (эндемичный род) и Hebe (Scrophula-riaceae), Celmisia, Olearia, Cotula, Abrotanella, Raoulia, Helichrysum и Senecio (Asteraceae), Astelia (Liliaceae), Luzula (Juncaceae), Uncinia и Carex (Cyperaceae).
По географическим связям виды новозеландской флоры очень разнородны. Основными элементами являются голантарктический (в широком смысле), палеоновозеландский, палеотропический (индо-малезийский, полинезийский и новокаледонский), австралийский, голарктический и космополитный. Как показывают данные палеоботаники, голантарктический элемент особенно процветал в позднем мелу и палеогене, когда Новая Зеландия была связана с Западной Антарктикой (Земля Мэри Бэрд). В это время были, в частности, хорошо представлены Araucaria (один вид которого сохранился только на о. Норфолк), Athrotaxis (в настоящее время только в Австралии и Тасмании), Gasuarina (полностью отсутствующая в Новозеландской области), а также более разнообразно представлены Not-hofugus и Proteaceae. Но уже в позднем мелу (около 80 млн. лет назад) Новая Зеландия начала отходить от Антарктики, и к миоцену ее северная часть вошла в зону субтропического климата. Палеотропический элемент постепенно стал заселять Новую Зеландию с севера через вулканические архипелаги, а в миоцене и позднее — вдоль ныне почти полностью погруженных хребтов Лорд-Хау и Норфолк. В результате в Новую Зеландию проникли такие тропические роды, как Macropiper, Metrosideros, Elaeocarpus, Homalanthus, Dysoxylum, Alectryon, Parsonsia, Avicennia, Elytranthe, пальмы, пандановое Freycinetia, многие орхидные. Ареал голантарктического элемента стал вновь расширяться в плиоцене и особенно в плейстоцене. В плейстоцене же проникли многие голарктические элементы.
Отделение Новой Зеландии от Антарктики и связанной с ней до среднего эоцена Австралии произошло еще до формирования характерной австралийской флоры, поэтому во флоре Новой Зеландии нет таких родов, как Acacia и Eucalyptus, а богато представленное в Австралии семейство Proteaceae имеет в Новой Зеландии только по одному виду родов Knightia (2 других вида в Новой Каледонии) и Persoonia (около 60 остальных видов в Австралии). Поэтому между Австралией и Тасманией и Новой Зеландией гораздо больше общих голантарктических элементов, чем собственно австралийских. С другой стороны, во флоре папоротников, которые древнее флоры цветковых растений, общий элемент между Новой Зеландией и Австралией и Тасманией достигает 40%.
Флористическое деление океана
Флористические области океана (цветная карта 3 на вклейке) совпадают с зоогеографи-ческими областями, установленными для морских животных. Наряду с водорослями это относится к морским цветковым растениям, которых насчитывается крайне ограниченное число родов и видов. В морях и океанах флористические области выделяются независимо для прикрепленных (бентосных) растений и свободноплавающих (планктонных) водорослей открытых пространств. Границы их часто не совпадают, так как гидрология и гидрохимия прибрежных и открытых вод могут быть различными; к тому же на распространение планктонных водорослей больше сказываются направление и сила течений. Для прикрепленных водорослей и морских цветковых растений большие открытые пространства океанов и морей часто служат непреодолимым препятствием.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://plant.geoman.ru/ 'Растения: виды, места произрастания, полезные свойства'