Бактериофаги и актинофаги
Во второй половине прошлого столетия учеными-микробиологами было открыто большинство бактерий, вызывающих заболевания человека, животных и растений. Были разработаны методы выделения этих бактерий и размножения их на искусственных питательных средах.
В то же время микробиологи обратили внимание на ряд инфекционных заболеваний человека и животных (оспа, бешенство, корь, краснуха и др.), а также растений, при которых бактериальные методы выяснения их возбудителей оказались непригодными.
Выходом из тупика, в котором очутилась микробиологическая наука, послужило открытие, сделанное русским ученым Д. И. Ивановским. В 1887 г. он, будучи еще студентом Петербургского университета, поехал на Украину и в Молдавию, где изучал причины мозаичной болезни табака, которая причиняла большой ущерб табачным плантациям южных районов. Свои исследования он проводил в течение нескольких лет и после окончания университета. В результате тщательного изучения этого заболевания Д. И. Ивановский показал, что оно вызывается специфическим возбудителем, состоящим из мельчайших частиц, или корпускул, которые по своим размерам значительно меньше бактерий.
Листья табака, пораженного мозаикой, он растирал в ступке и затем полученную массу фильтровал через специальные бактериальные фильтры, не пропускающие бактерий. Полученная после фильтрации прозрачная жидкость не содержала видимых в обычном микроскопе частиц. Но при нанесении ее на царапины, сделанные на поверхности здоровых листьев табака, растение заболевало мозаичной болезнью. Этот прием можно было повторять много раз.
Результаты исследований Д. И. Ивановского были опубликованы в 1892 г. в книге «О двух болезнях табака». В этой работе впервые в истории микробиологии было показано, что в природе существует особый мир мельчайших возбудителей инфекционных заболеваний. Эти возбудители проходят через бактериальные фильтры, невидимы в обычном микроскопе, не растут на тех средах, которые применяются для выращивания бактерий, и способны размножаться только в организме человека, животных и растений.
Эти мельчайшие организмы получили название фильтрующихся вирусов или вирусов.
Блестящее открытие Д. И. Ивановского ознаменовало новую эпоху в развитии микробиологии и заложило основы новой науки — вирусологии. Открытие первого вируса — возбудителя мозаики табака — было толчком к проведению широких исследований в области вирусологии.
Пользуясь предложенным Д. И. Ивановским методом, ученые стали открывать один за другим вирусы, вызывающие различные заболевания человека, животных и растений. В конце XIX — начале XX в. стало известно, что в природе существуют также вирусы, которые поражают бактерии и при этом вызывают распад (лизис) их клеток. Эти вирусы получили название бактериофагов — «пожирателей бактерий».
Явление лизиса, или распада, микробной клетки было известно давно. Оно может быть вызвано различными физическими и химическими факторами. В ряде случаев распад может происходить на определенном этапе развития микробной клетки как нормальный физиологический процесс ее старения. Однако имеется принципиальное различие между распадом клетки, вызванным бактериофагами, и химическими факторами. В первом случае происходит размножение бактериофага, вызывающего лизис, что не происходит при распаде клетки под влиянием физических и химических факторов.
Одним из первых, кто наблюдал и детально описал явление лизиса у бактерий, был один из основоположников отечественной медицинской микробиологии — Н. Ф. Гамалея. В 1896 —1898 гг. появились его работы, посвященные изучению явления лизиса у сибиреязвенной палочки. Фактор, вызвавший лизис этой бактерии, он назвал бактериолизином.
Для развития исследований в области бактериофагии особое значение имели работы французского ученого Д'Э р е л л я. В 1917 г. он сообщил, что из фекальных масс больных дизентерией ему удалось выделить особый лити-ческий фактор (вирус), способный проходить через бактериальные фильтры, размножаться на дизентерийных бактериях и вызывать при этом их лизис. Для обозначения этого вируса Д'Эрелль впервые предложил название бактериофаг. Кроме названия бактериофаг, или (сокращенно) фаг, в литературе, особенно в более старой, можно встретить также следующие: бактериофагический лизин, феномен Д'Эрелля, феномен Туорта, феномен Д'Эрел-ля — Туорта. Название «феномен Туорта» связано с именем английского микробиолога Туорта, который в 1915 г. описал явление перевиваемого лизиса у стафилококков и высказал предположение о вирусной природе этого явления.
Для обозначения фагов (вирусов микроорганизмов), вызывающих лизис актиномицетов, применяется термин актинофаг, микобакте-рий— микофаг, кишечной палочки — колифаг, водорослей — цианофаг и т. д.
Вначале много внимания уделялось изучению фагов, активных против патогенных бактерий: дизентерийной, брюшнотифозной, дифтерийной палочек, стафилококков с целью выяснения возможности использования их для лечения и профилактики инфекционных заболеваний. Одновременно много внимания уделялось изучению природы фагов.
Однако в последние годы проблема бактериофагии фактически превратилась в самостоятельную область биологии со своими специфическими разделами.
Фаги оказались весьма удобной моделью для разрешения ряда важнейших теоретических и практических вопросов общей биологии, генетики, молекулярной биологии, биохимии, а также медицины, ветеринарии и вирусологии.
Некоторые фаги отличаются специфичностью, т. е. они способны лизировать только определенные виды и разновидности бактерий. Поэтому такие фаги, названные типовыми, успешно применяются при дифференциации микроорганизмов. Созданы специальные музеи типовых фагов.
В результате большого теоретического и практического значения проблемы бактериофагии за последние 10—20 лет фаги изучались весьма интенсивно и всесторонне.
Морфология фагов
Применение современных электронных микроскопов, а также усовершенствование методов приготовления препаратов для электронной микроскопии позволили более детально изучить тонкую структуру фагов. Оказалось, что она весьма разнообразна и у многих фагов более сложна, чем структура вирусов растений и ряда вирусов человека и животных.
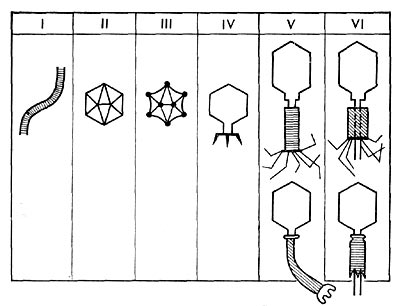
Рис. 204. Морфологические типы фагов.
Разные фаги отличаются друг от друга не только по форме, величине и сложности своей организации, но и по химическому составу. Оказалось, что фаги, лизирующие микроорганизмы различных групп, могут быть вполне идентичными по своей морфологии. В то же время фаги, активные против одной и той же культуры, могут резко различаться по своей структуре. Так, например, среди фагов, способных лизировать разные штаммы кишечной палочки, выявлены все известные морфологические типы фагов.
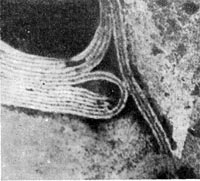
Рис. 205. Палочковидные, или нитевидные, фаги. Увел. X 400 000.
Частицы (или вирионы) большинства известных фагов имеют форму сперматозоида. Они состоят из головки (или капсида) и отростка. Наряду с этим есть фаги, которые состоят из одной головки, без отростка, и фаги, имеющие форму палочки (палочковидные или нитевидные фаги).
Рис. 206. Фаги второго морфологического типа, частица состоит из одной головки. Увел. X 600 000.
По форме частиц фаги делятся на шесть основных морфологических типов (групп): палочковидные или нитевидные фаги; фаги, состоящие из одной головки, без отростка; фаги, состоящие из головки, на которой имеется несколько небольших выступов; фаги, состоящие из головки и весьма короткого отростка; фаги, имеющие головку и длинный отросток, чехол которого не может сокращаться; фаги, имеющие головку и длинный отросток, чехол которого может сокращаться.
На рисунке 204 показано схематическое изображение частиц всех шести типов фагов. Размеры фагов принято обозначать в милли-микрометрах (1 миллимикрометр — миллионная часть миллиметра) или в ангстремах (10 А = 1 миллимикрометр).
Фаги первого морфологического типа — палочковидные или нитевидные — выявлены у кишечной, синегнойной, чудесной палочек и других бактерий. Средние размеры их: длина — от 7000 до 8500 А, ширина — от 50 до 80 А (рис. 205). Эти фаги отличаются от всех остальных не только большой специфичностью, но и рядом других важных свойств (о чем будет сказано дальше).
Риc. 207. Фаги третьего морфологического типа от головки отходят небольшие выступы. Увел. х 400 000.
Фаги второго морфологического типа. Частица их состоит из одной головки гексагональной (шестигранной) формы на плоскости. Частицы очень мелкие, средний размер их 230—300 А в диаметре (рис. 206).
У фагов третьего морфологического типа форма и размеры головки такие же, как у фагов второго типа, но у их головок имеются обычно несколько очень коротких выступов (рис. 207). Возможно, эти выступы являются аналогами отростков.
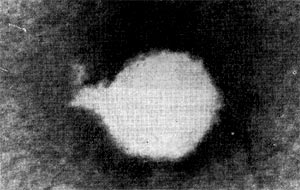
Рис. 208. Фаг четвертого морфологического типа. Частица состоит из головки ц короткого отростка Увел. х500 000.
Фаги второго и третьего морфологических типов отличаются постоянством формы и размеров, независимо от того, против каких микроорганизмов они активны. Эти фаги относятся к мелким формам.
Фаги четвертого морфологического типа. Частица состоит из головки, размеры которой варьируют от 400 до 640 А в диаметре, и очень короткого отростка (рис. 208). Длина и ширина отростка от 70 до 200 А.
Фаги пятого морфологического типа наиболее широко распространены. Головка у частиц гексагональной, формы различных размеров — от 500 до 4250 А в диаметре. Размеры отростка: длина — от 1700 до 5000 А, ширина — от 70 до 120 А (рис. 209). Чехол отростка не способен сокращаться.
Фаги шестого морфологического типа также широко распространены. Головка частицы разо-личной формы и размеров — от 600 до 1500 А в диаметре, гексагональная. Размеры отростка: длина — от 800 до 2890 А, ширина — от 140 до 370 А. Важной особенностью фагов этой группы является то, что чехол, окружающий отросток, способен сокращаться, в результате чего становится видимым внутренний стержень отростка (рис. 210).
Головки всех фагов состоят из внутреннего содержимого — нуклеиновой кислоты - - и окружены белковой оболочкой. Отросток фагов весьма сложен. Он обычно состоит из следующих структур: наружного чехла (или оболочки), внутреннего стержня с канальцем, базальной пластинки, оканчивающейся выступами (типа шипов) и нитевидными структурами. Чехол отростка состоит из субъединиц белковой природы, собранных в спираль. В результате этого он приобретает вид гофрированной трубки. В верхней части отростка многих фагов имеется образование, которое называется воротничком. На рисунке 211 схематически изображена тонкая структура фаговой частицы.
Химический состав фагов
Изучение химического состава фагов стало возможно лишь тогда, когда были усовершенствованы методы получения в больших количествах очищенных препаратов фага. В настоящее время изучен химический состав фагов, принадлежащих к разным морфологическим типам и поражающих микроорганизмы почти всех систематических групп.
Основными компонентами фагов являются белки и нуклеиновые кислоты. Важно отметить, что фаги, как и другие вирусы, содержат только один тип нуклеиновой кислоты — дез-оксирибонуклеиновую (ДНК) или рибонуклеиновую (РНК). Этим свойством вирусы отличаются от микроорганизмов, содержащих в клетках оба типа нуклеиновых кислот.
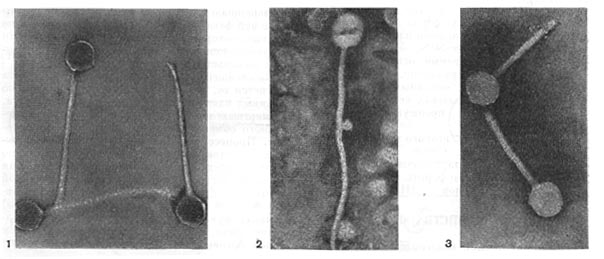
Рис. 209. Разные фаги пятого морфологического типа, частица состоит из головки и длинного отростка чехол которого не способен сокращаться. 1,2- увел. X 225 000, 3 - увел. X250 000
Нуклеиновая кислота находится в головке. Внутри головки фагов обнаружено также небольшое количество белка (около 3%).
Таким образом, по химическому составу фаги являются нуклеопротеидами. В зависимости от типа своей нуклеиновой кислоты фаги делятся на ДНК-овые и РНК-овые. Количество белка и нуклеиновой кислоты у разных фагов разное. У некоторых фагов содержание их почти одинаковое и каждый из этих компонентов составляет около 50%. У других фагов соотношение между этими основными компонентами может быть различно.
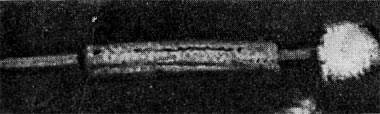
Рис. 210. Фаг шестого морфологического типа, частица состоит из головки и длинного отростка, чехол которого способен к сокращению. Увел, около 400 000.
Кроме указанных основных компонентов, фаги содержат в небольших количествах углеводы и некоторые преимущественно нейтральные жиры.
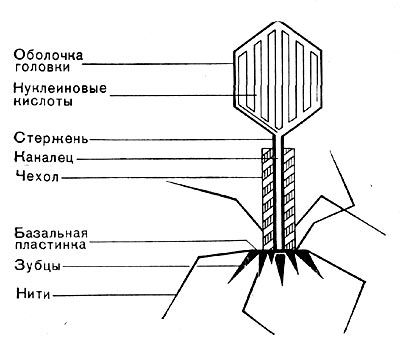
Рис. 211. Схема строения фаговой частицы.
Все известные фаги второго морфологического типа РНК-овые. Среди фагов третьего морфологического типа встречаются как РНК-овые, так и ДНК-овые формы. Фаги остальных морфологических типов — ДНК-овые.
Антигенные свойства фагов
Известно, что при введении в организм животного подкожно или внутривенно белка, бактериальных клеток, некоторых продуктов жизнедеятельности микроорганизмов и других веществ в крови животного вырабатываются вещества, названные антителами. Вещества, способные вызывать образование антител, называются антигенами.
Антитела очень специфичны и способны вступать в реакции только с теми антигенами, которые вызвали их образование. Они или связывают соответствующие антигены, или нейтрализуют их, или осаждают, или растворяют.
Оказалось, что все фаги обладают антигенными свойствами. При введении фага в организм животного в сыворотке крови образуются специфические антитела, способные действовать только против данного фага. Такие сыворотки называются антифаговыми. Когда фаг смешивается со специфической антифаговой сывороткой, происходит инактивация фага — фаг теряет способность вызывать лизис чувствительных к нему микробов.
Так как каждая антифаговая сыворотка специфична, ее можно успешно применять для идентификации и классификации фагов и очистки микробной культуры от фага. При помощи сыворотки удалось доказать, что белок оболочки фага отличается от белка оболочки отростка и от белка базальной пластинки и ее нитевидных образований, что говорит о сложности структуры фаговой частицы. По антигенным свойствам фаг резко отличается от чувствительных к нему микробов.
Размножение фагов
Взаимоотношения между фагом и чувствительной к нему клеткой очень сложны и не всегда завершаются лизисом клетки и размножением в ней фага. Мы рассмотрим такую инфекцию клетки, которая заканчивается гибелью клетки и размножением в ней фага. Такая инфекция называется продуктивной.
Важнейшей особенностью размножения фага является то, что оно может происходить только в живых клетках, находящихся в стадии роста.
В мертвых клетках, а также продуктах клеточного обмена размножение фага не происходит. Процесс размножения фага весьма сложный и состоит из следующих последовательно протекающих этапов (рис. 212): 1) адсорбция фаговой частицы на поверхности микробной клетки; 2) проникновение содержимого головки фаговой частицы (нуклеиновой кислоты) в микробную клетку; 3) внутриклеточное развитие фага, заканчивающееся образованием новых фаговых частиц; 4) лизис клетки и выход из нее новых фагов.
Время с момента инфицирования клетки фагом до лизиса клетки называется латентным или скрытым периодом. Продолжительность этого периода различна для разных типов фага, зависит от окружающей температуры, состава среды и других факторов. Латентный период фагов, специфичных для одних бактерий, 15—40 мин, для других — 5 ч и более. У фагов актиномицетов латентный период может быть еще продолжительнее. При низкой температуре латентный период значительно увеличивается.
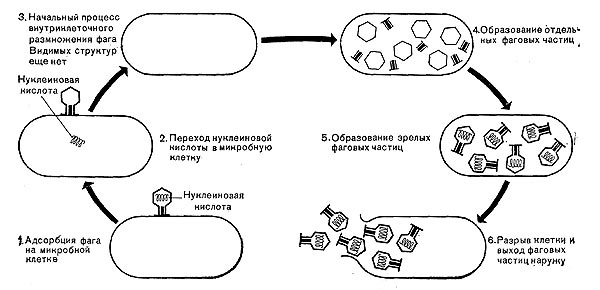
Рис. 212. Схема размножения фага.
Из всех этапов размножения фага наиболее изучен первый — адсорбция.
Адсорбция фага на клетке — реакция весьма специфичная. В клеточной стенке бактерий имеются особые структуры (рецепторы), к которым могут прикрепиться фаги. Адсорбируются на рецепторах только те фаги, к которым чувствительна клетка.
Фаги, имеющие отростки, прикрепляются к микробной стенке свободным концом отростка. Нитевидные фаги, а также фаги, не имеющие отростков, адсорбируются не на микробной стенке, а на нитевидных структурах, окружающих стенку, — фимбриях. Описаны фаги, которые прикрепляются отростком к бактериальным жгутикам. У некоторых фагов процесс адсорбции может осуществляться лишь в том случае, когда в среде имеются определенные вещества — кофакторы: аминокислоты (триптофан, тирозин и др.) или соли (кальциевые, магниевые).
На конце фагового отростка имеется особый фермент типа лизоцима. После адсорбции фага под влиянием этого фермента происходит растворение стенки микробной клетки и содержимое головки фага — нуклеиновая кислота — перекачивается в микробную клетку. Этим завершается второй этап процесса размножения фага.
Остальные структуры фаговой частицы — оболочка головки, отросток и его субструктуры — внутрь инфицированной фагом клетки не попадают. Их роль заключается в обеспечении сохранности фаговой частицы, находящейся вне клетки, и содействии проникновению фаговой нуклеиновой кислоты в клетку при инфекции.
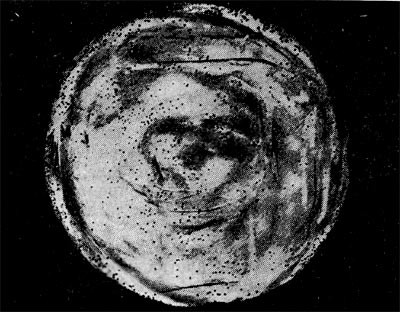
Рис. 213. Мелкие (едва видимые) негативные колонии актинофага. Увел. 6 : 10.
У нитевидных фагов, в отличие от других видов фагов, внутрь клетки проникает весь белок или его часть. После проникновения нуклеиновой кислоты фага в клетку начинается сложный процесс внутриклеточного размножения фага. Под влиянием нуклеиновой кислоты фага резко изменяется весь обмен микробной клетки. Основные процессы, протекающие в инфицированной клетке, направлены на образование новых фаговых частиц. Вначале формируются отдельно головки и отростки, которые затем объединяются в зрелые фаговые частицы. К этому времени внутри клетки образуется особый литический фермент, который вызывает лизис клетки изнутри. Клетка распадается, и новые зрелые частицы фага выходят наружу.
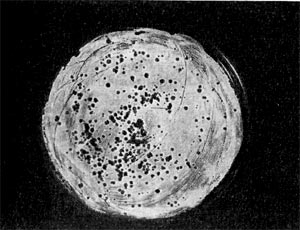
Рис. 214. Мелкие негативные колонии актинофага Увел. 8 : 9.
Количество новых фаговых частиц, образуемых одной клеткой при фаговой инфекции, называют выходом фага или его урожайностью. Выход фага зависит от свойств данного фага и не зависит от клетки-хозяина и ее размеров. Одни фаги отличаются очень низким выходом (5—50 частиц на клетку), у других выход значительно выше (от 1000 до 2500). Особенно высоким выходом отличаются мелкие РНК-овые фаги (свыше 20 000 частиц на клетку). Если большое количество бактериальных клеток смешать с небольшим количеством фаговых частиц, то процесс размножения фагов проходит несколько циклов. Вначале инфицируется часть клеток. Первое потомство фага инфицирует оставшиеся клетки — происходит второй цикл, за ним может следовать третий и т. д., пока не будут лизированы все чувствительные к данному фагу клетки. Среди фагов встречаются такие, размножение которых возможно лишь при наличии в среде определенных кофакторов. Одни из этих веществ, как уже указывалось, необходимы для адсорбции фага; другие — для внутриклеточного размножения фага.
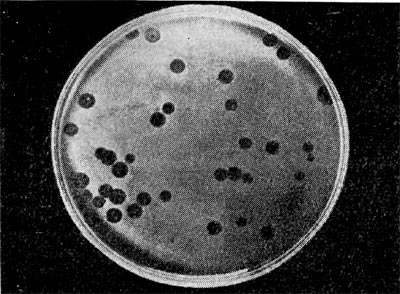
Рис. 215. Негативные колонии бактериофага средней величины. Увел. 1:1.
Является ли процесс размножения фага всегда смертельным для инфицированной им клетки? Несомненно, абсолютное большинство фагов вызывают при размножении лизис клетки и ее гибель. Лишь в последнее время было установлено, что при инфицировании клетки нитевидными фагами возможно размножение их без гибели клетки.
Если произвести рассев по поверхности ага-ризованной питательной среды в чашках Петри смеси фага и чувствительных к нему микробов и чашки выдержать в термостате, то происходит лизис клеток в результате размножения фага. Если взять большое количество частиц фага, то лизируется большая часть или весь выросший газон культуры. Если количество фаговых частиц таково, что они распределяются только на отдельных участках газона, лизируя в этих местах культуру, то возникает колония фага.
Эти колонии фага получили название бляшек, стерильных пятен. Правильнее их называть негативными колониями. Каждая негативная колония состоит из десятков и сотен миллионов фаговых частиц. Размер негативных колоний и их форма зависят в первую очередь от свойств фага, а также от состава среды и культуры микробов. У одних фагов негативные колонии очень мелкие и еле видимы невооруженным глазом, другие достигают 10 мм в диаметре и более. Колонии бывают светлые и четкие, когда лизировалась вся культура, или мутноватые, когда лизировались не все клетки. Вокруг негативных колоний некоторых фагов могут возникнуть различной формы и величины ореолы. На рисунках 213— 217 показаны негативные колонии разных фагов.
Морфология негативных колоний служит одним из признаков, которым пользуются при дифференциации фагов.
Распространение фагов
В настоящее время найдены, фаги, лизирую-щие клетки микроорганизмов, принадлежащих ко всем систематическим группам, как патогенных для человека, животных и растений, так и сапрофитных (непатогенных).
До недавнего времени не было ясно, существуют ли фаги против плесневых грибов и дрожжей. В последние годы найдены фаги, активные против грибов родов пенициллов, аспергил-лов и других, а также против некоторых дрожжей. Интересно отметить, что вирус удалось выявить и у тех видов пенициллов, которые применяются в промышленности для получения пенициллина. Не выявлены вирусы, активные против простейших животных, а также истинных спирохет.
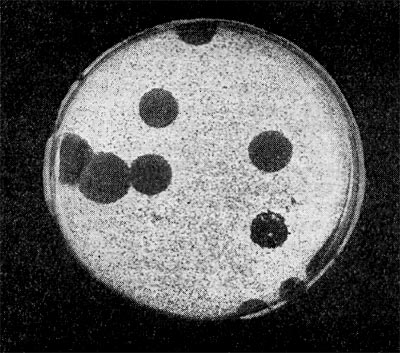
Рис. 216. Крупные негативные колонии актинофага. Увел. 1:1.
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, вода, выделения человека и животных и т. д.) микроорганизмами, тем в большем количестве в нем встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения. Фаги, активные против разных видов кишечной, дизентерийной, тифозной и паратифозной палочек, часто встречаются в содержимом кишечника человека и животных, сточных водах и загрязненных водоемах. Фаги фитопатогенных микроорганизмов успешнее всего выделяются из остатков растений, пораженных этими микробами.
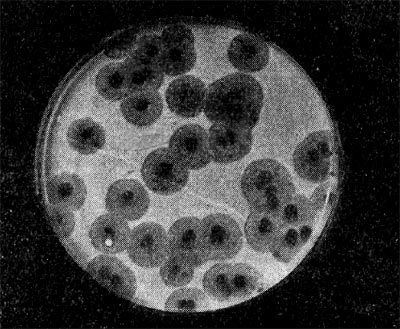
Рис. 217. Негативные колонии актинофага, окруженные ореолом угнетенного роста тест-культуры. Увел. 10 : 8.
Фаги молочнокислых стрептококков в большом количестве встречаются в молочных продуктах.
Итак, те субстраты, на которых развиваются определенные формы микроорганизмов, также благоприятны для существования соответствующих фагов.
Лизогения и её биологическое значение
При изучении явления бактериофагии исследователи обратили внимание на то, что иногда встречаются культуры микроорганизмов, которые содержат фаги, хотя на эти культуры фагами и не воздействовали. Явление фагоносительства получило название лизогении.
Оно было описано одним из основоположников учения о бактериофагах — Д'Эреллем, который считал, что такие культуры загрязняются фагом извне. Подобные культуры были названы ложнолизогенными.
Ложнолизогенные культуры состоят из смеси устойчивых и чувствительных к определенному фагу клеток.
Такие культуры могут быть легко освобождены от содержащихся в них фагов или путем нескольких рассевов, или с помощью специфической антифаговой сыворотки, или воздействием антифаговыми веществами.
Кроме ложнолизогенных, встречаются такие содержащие фаги культуры, у которых лизо-генное состояние, т. е. способность выделять фаги, стойко сохраняется даже после многочисленных пересевов в среде с антифаговой сывороткой и многократных воздействий антифаговыми веществами. Такие культуры названы истиннолизогенными. Мы расскажем только об этих культурах, которые будем называть лизогенными.
Лизогенными культурами являются такие культуры, которые обладают способностью продуцировать зрелые частицы фага без воздействия на них фагом извне. Это свойство стойко передается по наследству. В лизогенной культуре фаг находится внутри клетки. Для понимания сущности лизогении особо важное значение имел вопрос: в каком состоянии фаг находится внутри лизогенной клетки?
Опыты с искусственным разрывом клеток лизогенных культур под влиянием разнообразных физических и химических факторов (ультразвука, антибиотиков, литических ферментов и т. д.) не выявили наличия в клетках зрелых частиц фага.
Важное значение для понимания истинной природы лизогении имели работы А. Львова и А. Гутмана (1950). Оригинальные опыты этих исследователей убедительно показали, что в лизогенной клетке фаг находится не в виде зрелых частиц, а в какой-то другой, неинфекционной для клетки форме. Такую форму назвали профагом.
В зависимости от взаимоотношений фага и микроорганизма-хозяина фаги делят на две группы — вирулентные и умеренные, или сим-биотические. Вирулентные фаги вызывают лизис клетки. Умеренные фаги могут находиться в лизогенных клетках в виде профага.
Клетку можно экспериментально сделать лизогенной. Такой эксперимент помог выяснить механизмы процесса, благодаря которому клетка становится лизогенной. Оказалось, что при воздействии на клетку умеренным фагом часть популяции клеток лизируется, а другая часть становится лизогенной. При этом фаг адсорбируется клеткой и его нуклеиновая кислота проникает внутрь клетки. Однако, в отличие от продуктивной инфекции, вызываемой вирулентным фагом, при лизогенизадии нуклеиновая кислота фага связывается с ядерным аппаратом клетки (хромосомой) и остается в ней в виде профага.
Вопросы локализации профага в клетке и формы его связи с клеткой являются важнейшими проблемами лизогении.
По данным ряда исследователей, каждый профаг занимает определенное место на хромосоме лизогенной клетки. При делении клетки профаг воспроизводится со скоростью, равной скорости воспроизводства генетического материала клетки, что способствует передаче лизо-генного состояния потомству.
Следовательно, в лизогенной клетке профаг ведет себя как ее нормальный компонент. При лизогенизации происходит объединение генетического материала клетки с генетическим материалом фага на молекулярном уровне.
Известны пока единичные случаи, когда профаг не связан с хромосомой, а расположен на мембранах клеточной цитоплазмы.
Итак, в лизогенной клетке фаг является дополнительным генетическим фактором, который может неопределенно длительное время находиться внутри клетки и, как всякий генетический фактор, определять свойственные ему признаки.
Лизогенные культуры устойчивы (или иммун-ны) к тому фагу, который они содержат, а также к близкородственным ему фагам. При размножении лизогенной культуры какая-то часть клеток популяции лизируется и освобождает зрелые частицы специфичного для этой популяции умеренного фага.
Образование лизогенными культурами зрелых частиц фага получило название спонтанной индукции. Количество лизи-руемых клеток и количество образовавшихся зрелых частиц фага зависят от особенностей данной культуры и условий выращивания. В то же время количество клеток, освобождающих фаги, может быть резко увеличено при воздействии на лизогенную культуру некоторыми физическими и химическими факторами, получившими название индуцирующих. При индукции некоторых лизогенных культур удавалось вызывать образование зрелых частиц фага почти у всех клеток. К индуцирующим агентам относятся ультрафиолетовые (УФ), рентгеновские и гамма-излучения, перекиси, азотистый иприт и его гомологи, этиленимин, урацил, многие антибиотики. Наиболее эффективные и широко применяемые индуцирующие факторы — УФ-лучи и антибиотик митоми-цин С.
Как отмечалось, важным свойством лизогенной культуры является ее устойчивость к содержащемуся в ней фагу. В связи с этим выделение и изучение умеренных фагов лизогенной культуры возможно лишь в том случае, когда имеется другая культура того же вида, которая чувствительна к умеренному фагу данной лизогенной культуры. Такие культуры получили название индикаторных. К лизогенным культурам, особенно широко распространенным в природе, сравнительно легко можно подобрать индикаторные культуры среди других разновидностей этого же вида.
В отдельных случаях умеренный фаг лизогенной культуры может спонтанно (без внешних воздействий) или под влиянием различных факторов измениться и стать вирулентным. Тогда фаг приобретает способность лизировать все клетки данной культуры. У некоторых лизогенных культур превращение умеренного фага в вирулентный происходит сравнительно легко. Имеется ряд культур, у которых экспериментально не удавалось превратить умеренный фаг в вирулентный.
Возможность возникновения вирулентных мутантов умеренных фагов имеет большое теоретическое и практическое значение. Не редки случаи, когда единственным доказательством лизогенности культуры является возникновение вирулентных мутантов ее умеренного фага.
Лизогения широко распространена среди всех систематических групп микроорганизмов. Это явление детально изучено у сальмонелл — возбудителей брюшного тифа и паратифа, у дифтерийной палочки; все культуры этих видов патогенных бактерий оказались лизогенными. Лизогения широко распространена среди стрептококков, споровых форм бактерий, клубеньковых бактерий, актиномицетов, микобак-терий и др.; она выявлена и у некоторых мицелиальных грибов (пенициллов) и дрожжей.
Есть все основания утверждать, что абсолютное большинство микроорганизмов являются лизогенными. Ни про одну культуру нельзя с уверенностью сказать, что она не лизогенная.
За последнее время накапливается все больше данных о том, что многие лизогенные культуры содержат 2, 3, 4 и более умеренных фагов, т. е. являются полилизогенными. Например, многие актиномицеты, проактиномицеты, клубеньковые бактерии и некоторые спороносные бактерии содержат 4 и более фагов. Содержащиеся в полилизогенных культурах фаги часто резко различаются между собой по форме частиц, антигенным свойствам и спектру лити-ческого действия. Полилизогенные культуры можно экспериментально получить с помощью воздействия на них одновременно или последовательно различными умеренными фагами. Полученные таким способом культуры не отличаются от выделенных из природных источников.
Как уже отмечалось, профаг лизогенной культуры способен превратиться спонтанно или при индукции в зрелую полноценную фаговую частицу. Однако в ряде случаев под влиянием различных факторов у профага возникают стойкие наследуемые изменения (мутации), в результате которых он при индукции не способен превращаться в полноценную частицу. Поэтому у таких культур возникают частицы, состоящие только из головки или только из одного отростка. Возможны и другие нарушения в структуре фаговой частицы. При индукции таких культур лизогенная клетка лизируется, но образовавшиеся частицы как неполноценные не способны к размножению на индикаторной культуре.
Наиболее детально изучены дефектные фаги, у которых образуются одни лишь отростки. Такие фаги способны адсорбироваться на клетке, убить ее, но не могут размножаться. В последнее время такие дефектные фаги привлекли к себе внимание исследователей, так как было установлено, что многие описанные в литературе бактериоцины (вещества, убивающие бактерии) представляют собой дефектные фаговые частицы (табл. 59).
Существуют два принципиально различных типа бактериоцинов. Одни из них отличаются низким молекулярным весом, не осаждаются при центрифугировании, чувствительны к ферменту трипсину, термолабильны и в электронном микроскопе не видны. Бактериоцины другого типа обладают высоким молекулярным весом, осаждаются при центрифугировании, термостабильны и в электронном микроскопе видны в виде фагоподобных частиц или отдельных компонентов фаговой частицы (преимущественно в виде отростков).
О происхождении бактериоцинов первого типа и о возможной связи их с лизогенным состоянием культуры-продуцента никаких данных нет. В то же время многими исследователями показано, что образование бактериоцинов второго типа тесно связано с дефектной лизо-генией продуцента.
Наиболее убедительное доказательство дефектной лизогении —• выявление дефектных фаговых частиц, количество которых значительно увеличивается при индукции.
Имеются все основания утверждать, что дефектная лизогения довольно широко распространена. Она выявлена у очень многих культур, например у актиномицета, продуцирующего антибиотик стрептомицин (табл. 59, 1), клубеньковых бактерий (табл. 59, 2), спороносных бактерий, применяемых для борьбы с вредными насекомыми (табл. 59, 3). На рисунке 4 в табл. 59 показаны дефектные фаговые частицы, обнаруженные у кишечной палочки и названные колицином. Кроме того, были выявлены и описаны полилизогенные культуры актиномицетов, которые одновременно содержали нормальные и дефектные фаговые частицы (табл. 59, 5).
Как уже отмечалось, профаг в лизогенной культуре связан с ядерным аппаратом клетки и является дополнительным генетическим фактором. Профаг в лизогенной клетке ведет себя как ген, хотя между ними имеются принципиальные различия. За последние годы достигнуты большие успехи не только в изучении сущности лизогении, но и в выяснении роли профагов как дополнительных генетических факторов. Изменения, вызываемые профагом в лизогенной клетке, получили название л и-зогенных конверсии.
Лизогения, несомненно, одно из самых интересных явлений в биологии микроорганизмов, теоретическое и практическое значение которого далеко выходит за пределы микробиологии. Изучение этого явления сыграло большую роль в формировании представлений о фагах, их происхождении, о взаимоотношениях фагов с клеткой-хозяином.
Не исключено, что лизогенизация является одним из механизмов защиты микробной клетки от фаговой инфекции, выработанным клеткой в процессе длительной эволюции. Лизогенизация в известной степени биологически выгодна и клетке, и фагу. Клетка при лизогенизации становится устойчивой не только к данному фагу, но и к родственным ему фагам и, кроме того, приобретает дополнительные свойства.
Фаг же приобретает устойчивость к разнообразным внешним воздействиям и в то же время сохраняет потенциальную возможность перейти в вегетативное состояние и в состояние зрелой инфекционной частицы. Широкое распространение лизогении дает основание рассматривать это явление не как исключительное, а как нормальное на данном этапе эволюции микробов.
Изменчивость фагов и изменчмвость микроорганизмов под влиянием фагов
Фаги, как и микроорганизмы, способны изменять все свои свойства: форму и размеры негативных колоний, спектр литического действия, способность к адсорбции на микробной клетке, устойчивость к внешним воздействиям, антигенные свойства. Особенно часто наблюдаются изменения морфологии негативных колоний, спектра литического действия и превращение умеренных фагов в вирулентные.
Большие изменения могут наблюдаться в тонкой структуре фаговой частицы — возникают дефектные частицы, лишенные головки, отростка, нитевидных образований или других субструктур.
Изменения фагов могут быть наследственными (мутации) и неиаследственными (феноти-пические). Фенотипические изменения зависят от условий, в которых образуются фаговые частицы. Важное значение имеют изменения, вызываемые клеткой-хозяином, т. е. той культурой, на которой фаг размножается. Эти изменения большей частью носят фенотипический характер и касаются преимущественно формы негативных колоний, спектра литического действия и вирулентности. Под влиянием клетки-хозяина возможны и стойкие изменения типа мутаций. С помощью разных мутагенных факторов (лучистой энергии, химических агентов) могут быть получены разнообразные мутанты.
Особый интерес представляют изменения, происходящие при одновременном размножении на одной и той же культуре двух родственных по антигенным свойствам фагов. При этом в потомстве возникают частицы каждого из этих фагов и, кроме того, формы, которые приобрели свойства обоих родителей (гибридные формы).
Под влиянием фагов могут существенно изменяться все свойства микроорганизмов — морфология клеток, строение колоний, токсичность, подвижность и т. д. Изменения, вызываемые фагом, могут быть наследственными и ненаследственными. Механизмы, приводящие к изменению клеток под влиянием фагов, различные. В отдельных случаях фаг играет лишь роль отбирающего фактора: под его влиянием лизируются все чувствительные к нему клетки данной популяции и остаются лишь те клетки, которые еще до воздействия фагом были по разным причинам устойчивы к нему.
При выращивании микроорганизмов в жидкой среде совместно с активным против них фагом обычно наблюдается следующее. Сначала среда мутнеет, а затем просветляется в результате лизиса клеток. В ряде случаев через некоторое время (разное для разных микробов и фагов) культуральная жидкость снова мутнеет. Помутнение среды происходит вследствие возобновления роста культуры. На агаризованных средах при нанесении фага на газон чувствительной к нему культуры можно наблюдать вначале лизис культуры в местах нанесения фага, а через некоторое время на лизированных участках появляется рост культуры, обычно в виде отдельных колоний (рис. 218).
Вырастающие после лизиса микроорганизмы получили название культур вторичного роста. Анализ культур вторичного роста показывает, что в одних случаях они состоят из вариантов, ставших стабильно устойчивыми к данному фагу, в других — из нестойких форм. Культуры, ставшие устойчивыми к фагу, могут одновременно приобрести и ряд новых свойств. Этим обычно пользуются при получении фагоустойчивых культур для промышленных целей.
В последние годы были выявлены два принципиально различных механизма изменчивости клеток под влиянием фагов — трансдукция и лизогенные конверсии.
Трансдукция (перенос)
При размножении определенных умеренных фагов на чувствительних к ним культурах фаговая частица захватывает какой-нибудь фрагмент генетического материала данной клетки. При воздействии этим же фагом на другую чувствительную к нему культуру он передает новой культуре захваченный фрагмент. Культура, от которой фаг переносит генетический материал, получила название донора (дающая), а культура, приобретающая генетический материал, — реципиента (получающая).
При трансдукции фаг играет роль механического переносчика; лизогенизация клетки не обязательна. Один и тот же фаг может переносить разные свойства.
Трансдукция происходит довольно редко: из одного и более миллионов фаговых частиц только одна способна осуществлять трансдук-цию. При помощи трансдукции удавалось перенести от клеток-доноров клеткам-реципиентам различные свойства: токсичность, устойчивость к антибиотикам, способность продуцировать определенные ферменты, антигенные и другие свойства.
Лизогенные конверсии (превращения).
Как уже отмечалось, при лизогенизации клетка-хозяин приобретает устойчивость к данному фагу, а также способность продуцировать зрелые частицы этого фага. Однако этим не ограничиваются изменения, вызванные фагом при лизогенизации. Многочисленными опытами на микроорганизмах самых различных систематических групп было показано, что при лизогенизации клетка приобретает новые, точно определенные свойства, характер которых зависит от особенностей данного фага.
В отличие от трансдукции, при которой фаг выступает в роли механического переносчика генетического материала, при лизогенизации сам фаг (вернее, его нуклеиновая кислота) является тем генетическим материалом, который в виде профага придается генетическому материалу клетки. Поэтому при лизогенизации не имеет значения культура, на которой размножался данный умеренный фаг.
Наиболее детально лизогенные конверсии изучены у некоторых патогенных бактерий, преимущественно у дифтерийной палочки и сальмонелл.
Дифтерийная палочка содержит три разных фага. Оказалось, что только один из них (фаг бета) влияет на образование этой культурой токсина. При отсутствии в клетке фага бета культура не продуцирует токсина. Если нетоксичную дифтерийную культуру лизогенизиро-вать фагом бета, то она приобретает способность образовывать токсин.
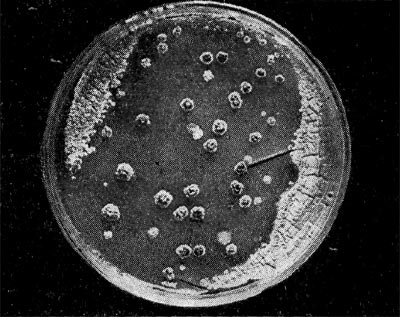
Рис. 218. Появление колоний вторичного роста после лизиса актинофагом. Культура Act. oriental's продуцирует антибиотик ванкомицин.
Опытами на сальмонеллах было показано, что токсичность, антигенный свойства, подвижность и другие признаки связаны с наличием в этих культурах строго определенных фагов. Среди споровых бактерий группы клостридий имеются виды, образующие ботулинический токсин, вызывающий опасные пищевые отравления. Недавно удалось выяснить, что эти культуры полилизогенные и один из содержащихся в них фагов вызывает образование токсина. Для ряда культур показано, что способность продуцировать определенные ферменты, антигенные и другие их свойства связаны с лизогенизацией определенными фагами. Была выявлена одна полилизогенная культура акти-номицета, которая содержала 4 разных фага. Из них один определял способность культуры продуцировать антибиотик. После удаления его культура теряла способность продуцировать антибиотик. При лизогенизации неактивной культуры этим фагом она вновь приобретала способность образовывать антибиотик.
К сожалению, еще у очень многих лизоген-ных культур микроорганизмов не установлено, какие свойства их связаны с фагами. Выяснение этого вопроса — одна из важнейших задач микробиологов и генетиков.
Природа фагов
Несмотря на то что явление бактериофагии интенсивно изучается более пятидесяти лет, на природу фагов нет единой точки зрения и этот вопрос до сих пор остается спорным. В нашей стране и за рубежом в специальной печати и на конференциях неоднократно возникали горячие дискуссии на эту тему. И это не случайно. Вопрос о происхождении фагов, как и других вирусов, имеет большое значение, так как с ним тесно связано решение многих актуальнейших задач современной биологии: происхождение жизни, возможные формы существования живого; существование живых существ, не имеющих клеточной структуры; происхождение клеточных форм жизни; развитие, изменчивость и видообразование у микроорганизмов и др.
До настоящего времени все еще существуют диаметрально противоположные точки зрения на природу вирусов, в том числе и фагов. По мнению одних ученых, фаги относятся к живым организмам; другие рассматривают их как особые вещества типа ферментов.
Важно отметить, что те исследователи, которые относят фаги к живым организмам, различно трактуют вопрос об их происхождении. Одни исследователи считают, что фаги, как и вирусы человека, животных и растений, произошли от древнейших доклеточных форм, которые в процессе эволюции приспособились к паразитированию в первичных одноклеточных организмах и в дальнейшем эволюционировали вместе со своими хозяевами. Таким путем, как думают эти ученые, возникли фаги микроорганизмов, которые по своему отношению к клетке-хозяину являются паразитами экзогенного, т. е. внешнего, происхождения. Другие же считают, что происхождение фагов связано тем или иным образом с клеткой своего теперешнего хозяина (эндогенное происхождение).
По мнению ученых, рассматривающих фаг как фермент эндогенного происхождения, фаговая частица является продуктом жизнедеятельности микробной клетки. При попадании в клетку фаги вызывают каталитически протекающие процессы образования активного фага, способного разрушать микробную клетку. А размножение фага в клетке происходит приблизительно так же, как образование активного фермента из его неактивного предшественника — профермента.
Какая же из изложенных точек зрения на природу фага является наиболее приемлемой, исходя из современных знаний о свойствах фага и его взаимоотношений с клеткой-хозяином? Является ли фаг живым существом или это — вещество подобное ферменту? За последние годы благодаря применению новейших современных методов исследования (электронная микроскопия, меченые атомы) знания о структуре фагов, их химическом составе, особенностях размножения значительно расширились. Фаговая частица оказалась довольно сложно организованной. Она содержит основные химические соединения, свойственные живому организму,— нуклеиновые кислоты и белок. Подобно другим живым существам, фаги способны изменять все свои свойства. Поэтому рассматривать их как ферменты нет никаких оснований. Верно, они не обладают собственным обменом веществ. Они являются абсолютными паразитами, живущими полностью за счет клетки-хозяина.
Следовательно, фаги логично рассматривать как особые формы живых существ. Однако вопрос о происхождении фагов пока еще нельзя считать окончательно решенным.
Проблема фагии в различных производствах
За последние 30—40 лет значительно увеличилось количество производств, основанных на использовании продуктов жизнедеятельности микроорганизмов. При помощи микроорганизмов получают разнообразные антибиотики, ферменты, витамины, аминокислоты, ацетон и бутиловый спирт, молочную, уксусную, лимонную и другие кислоты, различные бактериальные препараты — удобрения, инсектициды и многие другие.
Вследствие концентрации на указанных производствах больших масс микроорганизмов, находящихся в стадии интенсивного роста, создаются благоприятные условия для размножения соответствующих фагов, часто вызывающих лизис производственных культур. Впервые с этим явлением столкнулись в сыродельном производстве, для которого борьба с фаголизисом — одна из актуальнейших проблем.
Микробиологические производства, которые сталкиваются с проблемой фагии, можно разбить на две группы. К первой группе относятся те производства, в которых основное применяемое сырье может содержать (и большей частью фактически содержит) фаги. Это предприятия молочной промышленности. Одно из древнейших производств — молочное (сыроделие, получение молочнокислых продуктов и др.) — основано на применении молочнокислых стрептококков и бактерий.
Ко второй группе производств относятся те, в которых выращивание микроорганизмов производится на питательных средах, не содержащих фагов. Это фактически все остальные микробиологические производства.
Из производств первой группы рассмотрим сыроделие как наиболее типичное и в то же время лучше изученное с точки зрения фагии.
Молочнокислые микробы очень широко распространены в природе: в почве, в навозе, на корнях, листьях и семенах растений, в кишечнике животных. Широкому распространению фагов молочнокислых бактерий способствует то, что они являются лизогенными, т. е. содержат внутри клеток фаги. В результате этого в молоке сразу же после доения уже есть фаги. Молоко, поступающее на молочные заводы, часто содержит фаги в значительном количестве (1 млн. и более частиц в 1 мл молока). Следует отметить, что все фаги, даже активные против патогенных микробов, совершенно безвредны для людей и животных.
Современное производство сыра основано на применении пастеризованного молока. Однако пастеризация не убивает всех имеющихся в молоке фагов. Охлажденное после пастеризации молоко разливают в специальные ванны, в которые вносится закваска, состоящая из чистых культур молочнокислых стрептококков. Стрептококки вызывают сквашивание молока. Получаемый сгусток молока подвергается дальнейшей переработке.
Работники сыродельных заводов давно обратили внимание на то, что в ряде случаев активность молочнокислых микробов закваски резко снижается, что приводит к плохому сбраживанию молока. Это явление может быть вызвано разными причинами. Но чаще всего оно вызывается фагами, которые лизируют полностью или частично культуры заквасок. В результате этого процесс молочнокислого брожения полностью останавливается или интенсивность его резко снижается.
На сыродельных заводах, как правило, применяют закваски, состоящие не из одной культуры, а из смеси различных культур молочнокислых стрептококков.Стрептококков очень много в природе, поэтому для производства можно отобрать культуры, отличающиеся по своей чувствительности к фагам. При применении смешанной закваски под влиянием фага лизи-руется одна или две культуры, другие же продолжают процесс молочнокислого брожения.
Одну и ту же закваску используют только определенное время, после чего ее заменяют другой. Длительно применять одну и ту же закваску нельзя, так как это способствует накоплению на заводе фагов, активных против культур данной закваски.
Правильный выбор культур для заквасок, смена их на основании изучения появившихся на заводе фагов дают значительный эффект. В ряде зарубежных стран на сыродельных заводах применяют закваски, содержащие только одну культуру. В этих случаях одну и ту же закваску используют лишь один раз.
Успешность борьбы с фаголизисом требует проведения и ряда других мероприятий. Особое значение имеет борьба с распространением фагов в заводских помещениях.
Значительные трудности в связи с лизисом под влиянием фагов производственных культур испытывала у нас и за рубежом антибиотическая промышленность.
Большой ущерб наносят фаги заводам аце-тонобутиловым, а также изготавливающим бактериальные удобрительные препараты и препараты, применяемые для борьбы с вредными насекомыми.
Возникает вопрос: как фаги попадают на эти заводы? Есть два источника. Первый и, по-видимому, основной для антибиотической промышленности — это сами культуры, применяемые для получения антибиотиков. Все эти культуры лизогенные и содержат внутри своих клеток умеренные фаги. Сами умеренные фаги не опасны. Но в ряде случаев они могут измениться и стать вирулентными, т. е. способными лидировать культуру хозяина. Вирулентные фаги, накапливаясь на заводах, вызывают массовый лизис производственных культур.
Другим источником попадания фагов на производства является почва, которая содержит много фагов. Вместе с пылью фаги могут заноситься в заводские помещения и заражать производственную культуру.
В борьбе с фаголизисом, в особенности при производстве антибиотиков, большое значение имело экспериментальное получение активных фагоустойчивых культур. Такие культуры в настоящее время успешно применяются.
Практическое использование фагов
Первыми были выделены фаги, активные против патогенных микроорганизмов (дизентерийной палочки).
Вполне естественно, что у исследователей многих стран возник вопрос об использовании фагов для лечения и профилактики инфекционных заболеваний, вызываемых бактериями (дизентерия, брюшной тиф, паратифы, холера и др.). Многочисленные исследования в этой области были проведены и в нашей стране. Результаты этих работ весьма разноречивы.
Имеются данные, показывающие несомненную эффективность фагов при лечении дизентерии и холеры. Во время Великой Отечественной войны некоторые хирурги успешно применяли фаги для борьбы с нагноением ран.
Вместе с этим накопились многочисленные данные об отсутствии лечебного эффекта при применении фагов. Одна из основных причин низкой эффективности или полного отсутствия лечебного эффекта заключается в неумелом подборе фагов для лечебных целей. Одна и та же болезнь, например дизентерия, может вызываться различными видами Л разновидностями дизентерийных бактерий. Фаги, активные против одних дизентерийных бактерий, совершенно не влияют на другие. Это не всегда учитывалось в должной мере при приготовлении фаговых препаратов для лечения определенных заболеваний.
В последние годы фаги для лечебных целей почти не применяются. Определенное влияние на отрицательное отношение к использованию фагов в лечебных целях сыграло не только непостоянство результатов, но и появление многочисленных антибиотиков, а также разных хемотерапевтических препаратов.
Совершенно иначе обстоит дело с использованием фагов для профилактических целей. Убедительно показано, что при появлении в детских садах, яслях, школах дизентерийной инфекции использование препаратов фага предотвращает заболевание детей, имевших контакт с заболевшими. В настоящее время готовят сухие препараты фагов в виде драже.
Среди фагов встречаются такие, которые отличаются специфичностью, т. е. способны лизировать культуры только определенного вида или разновидности. Такие фаги, получившие название типовых, успешно применяются при классификации культур. Есть специальные коллекции типовых фагов, активных против патогенных микроорганизмов. Эти фаги помогли выяснить источники многих заболеваний. При помощи специфических фагов можно установить наличие определенных патогенных и непатогенных форм микробов в воде и в выделениях кишечника, а также наличие фитопатогенных бактерий внутри семян растений.
Из лизогенных культур актиномицетов выделен ряд актинофагов, которые можно использовать при классификации актиномицетов. По чувствительности культуры актиномицета к определенному фагу можно было судить, какой она продуцирует антибиотик.
В последние годы было установлено, что способностью индуцировать лизогенные культуры обладают многие вещества с противоопухолевым действием. Поэтому лизогенные культуры стали успешно применяться в поисках антираковых веществ.
Фаги оказались весьма ценной, а в ряде случаев и незаменимой моделью для решения многих вопросов молекулярной биологии, генетики, общей вирусологии и онкологии.
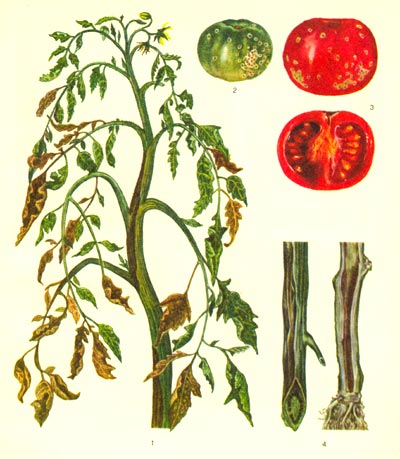
Таблица 58. Бактериальный рак томатов: 1 - больной куст; 2 - зелёный больной плод, пятна 'птичий глаз'; 3 - больной спелый плод и его поперечный разрез; 4 - поперечный и продольный разрезы больного стебля (побурение сосудов).
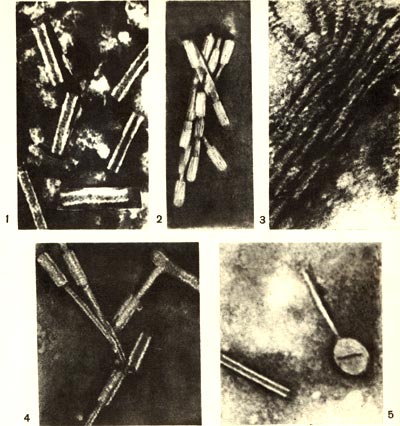
Таблица 59. Дефектные фаговые частицы: 1—Actinomyces streptomycini — продуцента антибиотика стрептомицина; 2 — клубеньковых бактерий люцерны; 3 — энтомопатогенных бацилл; 4 — кишечной палочки; 5— полилизогенная культура актиномицета, содержащая нормальные и дефектные фаговые частицы. Увел.: / -- х 400000; 2-х 160000; 3-х 520000; 4 — х 100000: 5 —х 300 000. 1—Actinomyces streptomycini — продуцента антибиотика стрептомицина; 2 — клубеньковых бактерий люцерны; 3 — энтомопатогенных бацилл; 4 — кишечной палочки; 5— полилизогенная культура актиномицета, содержащая нормальные и дефектные фаговые частицы. Увел.: / -- х 400000; 2-х 160000; 3-х 520000; 4 — х 100000: 5 —х 300 000.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://plant.geoman.ru/ 'Растения: виды, места произрастания, полезные свойства'