Термофильные бактерии
Экологически обособленную группу в природе представляют термофильные микроорганизмы. Температурные условия вызывали в в процессе эволюции появление микробных форм, которые оказались способными развиваться при разных температурах, в том числе и при высокой (50—93 °С).
Видная роль в изучении термофильных микроорганизмов принадлежит А. А. Имшенецкому, Е. Н. Мишустину, Б. Л. Исаченко и др. Эти ученые не ограничились разработкой только теоретической стороны проблемы явления термофилии, и их исследования имели важное практическое значение.
Одна из главных отличительных особенностей термофилов — ускоренный обмен веществ. За последние годы благодаря новейшим методам исследования удалось накопить данные, частично раскрывающие механизмы, при помощи которых клетка защищается от воздействия высокой температуры. Установлено, что наиболее существенные изменения под воздействием высокой температуры претерпевают клеточные белки и липиды, с которыми связаны основные жизненные процессы.
Благодаря высокой скорости роста термофильные микроорганизмы могут найти широкое применение в самых различных отраслях промышленности и сельского хозяйства.
Методы выделения термофильных и мезофиль-ных бактерий в основном сходны. Различие заключается лишь в температуре выращивания. Для того чтобы точно установить оптимальную температуру развития и закрепить ее, культуру необходимо длительно (1 — 2 месяца) пассировать (пересевать) в диапазоне найденного оптимума.
Температурные группы микроорганизмов. Термртолератность и термоустойчивость
Различные микроорганизмы могут развиваться при разных температурах: одни микробы хорошо растут при низких температурах, близких к О °С (+5 °С); другие, наоборот, способны к росту при высоких температурах (около 90 °С). Поэтому микроорганизмы делят по их отношению к температуре на три основные группы— психрофилы, мезофилы и термофилы.
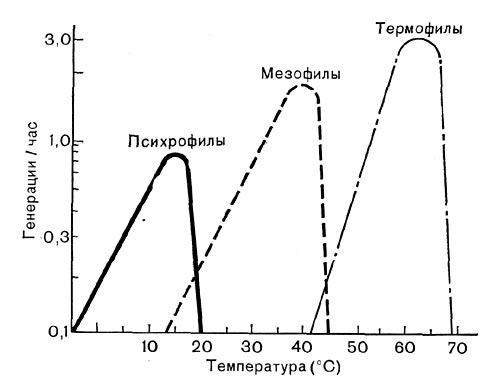
Рис. 187. Зависимость от температуры скорости роста психрофильных, мезофильных и термофильных микроорганизмов.
Психрофилы (предпочитающие низкие температуры) — микроорганизмы, имеющие минимальную температуру роста ниже О °С.
Мезофилы (предпочитающие средние температуры) — микроорганизмы, имеющие минимальную температуру роста выше, чем у психрофилов, а максимальную температуру ниже, чем у термофилов. Большинство микроорганизмов — мезофилы, растущие обычно при температурах от 0—10 °С до 40—45 °С.
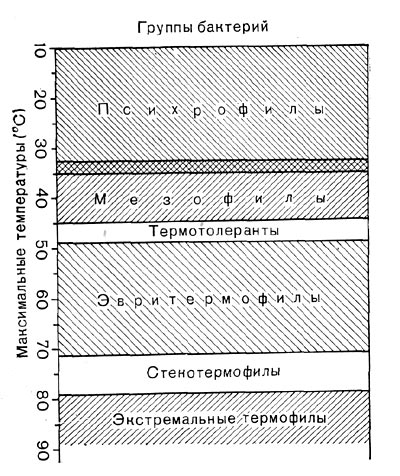
Рис. 188. Группы бактерий по максимальным температурам роста.
Термофилы (предпочитающие высокие температуры) — микроорганизмы с максимальной температурой роста обычно выше 50 °С.
Что же такое минимальная и максимальная температуры роста? Минимальная — это такая пороговая температура, при незначительном снижении которой скорость роста микроорганизма (прирост клеток за 1 ч) близка к нулю, т. е. практически рост прекращается. Максимальная температура — пороговая температура, при незначительном повышении которой скорость роста микроорганизма близка к нулю.
При изучении особенностей каждого нового штамма обязательно определяют и оптимальную температуру, т. е. устанавливают тот температурный диапазон, в котором данный микроорганизм растет с максимальной скоростью. При максимальной скорости роста микроорганизм, естественно, наиболее интенсивно размножается. Поэтому довольно часто скорость роста выражают как число генераций за 1 ч. Английский ученый Т. Д. Брок предложил схему, показывающую зависимость скорости роста (частоты генераций) от температуры для различных температурных групп микроорганизмов (рис. 187).
Внутри группы термофилов различают еще четыре более мелкие подгруппы (рис. 188) — экстремально термофильные микроорганизмы, стенотермофилы, эвритермофилы, термотолерантные микроорганизмы (термотолеранты).
Экстремально термофильные микроорганизмы вовсе не растут при температурах ниже 40—45 °С, оптимальная температурная зона роста — около 80 °С, максимальные температуры близки к 93 °С.
Стенотермофилы характеризуются минимальными температурами роста, равными 37—40 °С, максимальные лежат в области 70— 80 °С, зона оптимальных температур — 55— 65 °С.
Наибольшее количество термофильных микроорганизмов относится к подгруппам эвритер-мофилов и термотолерантов. Эти подгруппы довольно сложно четко охарактеризовать. Особенно трудно отличить термотолерантные штаммы от некоторых мезофилов.
Эвритермофилы имеют минимальную температуру роста ниже 37 °С, а максимальную — выше 48 °С, но ниже 70 °С. Эта подгруппа включает представителей различных систематических групп — бактерий, акти-номицетов, дрожжей, грибов, водорослей.
Термотолеранты характеризуются максимальной температурой роста, равной 45—48 °С (для бактерий). Однако некоторые мезофильные штаммы также могут иметь максимальную температуру, равную 45 °С. В таких случаях отличить термотолерантный штамм от мезофильного можно по изменению величины скорости роста при незначительном (на 3—6°) сдвиге температуры в сторону возрастания от значения температур, являющихся оптимальными для мезофильных штаммов бактерий (обычно 37 °С). При таком увеличении температуры скорость роста термотолерантного микроорганизма существенно не изменится, а мезо-фильный штамм будет развиваться с заметно снижающейся скоростью. Если микроорганизм окажется эвритермофильным (со сравнительно низкой максимальной температурой 48—50 °С), то его скорость роста при повышении температуры от 37 до 43 °С резко возрастет.
Таким образом, по изменению скорости роста можно установить принадлежность данного штамма микроорганизма к той или иной температурной группе или подгруппе.
Итак, термотолерантный микроорганизм способен размножаться с почти одинаковой скоростью как при обычной температуре (37 °С), являющейся оптимальной для мезофильных штаммов, так и при более высокой (на 3—7° выше) температуре. Термотолерантный микроорганизм как бы безразличен к такому изменению температуры. Да и сам термин «термотолерантность» означает терпимость к повышению температуры. Следовательно, под термотолерантностью следует понимать способность микроорганизма размножаться без существенного уменьшения или увеличения скорости роста при указанном выше увеличении температуры. В природе и лабораторных условиях микроорганизмы могут подвергаться кратковременному воздействию высоких температур. Во время такого теплового воздействия клетки обычно не размножаются. После прекращения действия этого неблагоприятного для развития микроорганизма фактора одни штаммы могут сохранить репродуктивную способность (способность к размножению), другие оказываются менее устойчивыми и погибают. Устойчивость микроорганизмов различных температурных групп (психрофилов, мезо-филов, термотолерантов, термофилов) к кратковременному воздействию высоких температур без повреждения репродуктивной способности микроорганизма (при снятии действия этих температур) целесообразно характеризовать термином термоустойчивость (термо-резистентность).
Причины, обуславливающие способность термофилов существовать при высоких температурах
Многие ученые давно пытались установить, почему термофильные микроорганизмы могут существовать при таких высоких температурах, как 50—90 °С. Оказалось, что как структурные и клеточные элементы, такие, как оболочка, мембраны, рибосомы, так и входящие в состав клетки протеины, жиры, ферменты заметно отличаются качественно и количественно от подобных клеточных компонентов мезофильных форм.
Выяснилось также, что если некоторые элементы клетки недостаточно стабильны к высокой температуре (например, рибонуклеиновые кислоты), то их синтез в клетке осуществляется с большей скоростью. В этом процессе участвуют ферменты, отличающиеся высокой термостабильностью.
Оболочка клетки термофилов также обладает заметной устойчивостью к действию температуры. Это обусловлено ее химическим составом и более устойчивым к температуре механизмом (чем у мезофилов), осуществляющим синтез клеточных стенок.
Д ж. Т. Форрестер и А. Д ж. Вике н показали, что содержание липидов в клеточных стенках термофилов выше, чем в стенках большинства мезофилов. В клеточных стенках Вас. coagulans, выращенных при 55 °С, обнаружен необычный полимер, подобный тейхоевой кислоте.
Значительный интерес представляют ранние исследования X. Коффлера, который показал, что клеточные белки, выделенные из жгутиков термофилов, более термостабильны, чем белки, выделенные из жгутиков мезофильных форм микробов.
Американский исследователь Т. Д. Брок высказал мнение о решающей роли клеточной мембраны в предохранении клетки от теплового повреждения. Некоторые исследователи выдвигают предположение, что состав мембранных липидов определяется максимальной или минимальной температурой роста микроорганизмов. Так, показано, что среди мембранных липидов Bacillus stearothermophilus преобладают жирные кислоты с более длинными и разветвленными цепочками (С15 и С17). Вероятно, эти кислоты могут придавать большую упругость мембранной структуре.
Помимо качественных различий в химическом составе клеточных мембран, клетки термофильных бактерий содержат больше мембран, чем клетки мезофильных бактерий. X. Бодман и Н. Е. Велкер нашли увеличение количества мембран в клетках Вас. stearothermophilus при повышении температуры роста. Так, при температуре роста Вас. stearothermophilus 55 °С на долю мембран приходилось 16,5 %, а при 65 °С — 17,8% от сухой массы клеток. Также было отмечено, что с повышением температуры роста указанной культуры от 55 до 65 °С отношение протеинов к липидам в мембранах возрастает от 3,65 до 5,22 соответственно.
Клеточные мембраны могут иметь различное строение у разных групп микроорганизмов. Так, например, известно, что у грамотрицатель-ных бактерий мезосомы менее развиты, чем у грамположительных. Возможно, поэтому основные виды облигатных термофильных бактерий грамположительны.
Мембраны в клетке служат не только границей раздела фаз, но и местом локализации ряда важнейших биологических систем. В настоящее время имеются данные, указывающие, что некоторые ферменты термофилов приобретают термостабильность, когда они связаны с клеточной мембраной. Если их отделить от мембраны с помощью ультразвука, то эти ферменты становятся относительно термолабильными.
Мембраны термофильных бактерий отличаются высокой механической прочностью.
Было показано, что многие облигатно-термо-фильные бактерии (Вас. stearothermophilus, Вас. coagulans, Вас. denitrificans) способны образовывать шаровидные протопласты при развитии сначала в интенсивно аэрируемой среде, а затем (в фазе экспоненциального роста) без аэрации.
Одним из главных факторов, вызывающих процесс образования протопластов, является резкий дефицит кислорода в среде. Образовавшиеся в этих условиях протопласты могут длительное время не лизироваться в обычных условиях, что свидетельствует об их значительной осмотической устойчивости.
Рибосомы, выделенные из клеток облигатно-термофильных бактерий, обладают значительно большей термостабильностью, чем рибосомы, выделенные из клеток мезофильных форм. Высказывается предположение, что наибольшая стабильность рибосом у термофилов может быть благодаря различиям в составе и структуре рибосомальных протеинов.
Интересно привести данные, полученные при изучении термостабильности рибосом у Thermus aquaticus, оптимальная температура роста которой равна 70 °С.
Рибосомы грамотрицательной палочки Thermus aquaticus были устойчивы при нагревании до 79 °С, а рибосомы Bacterium coli разрушались при увеличении температуры до 59 °С. Температура денатурации рибосом Т. aquaticus коррелирует с максимальной температурой роста этой бактерии, также равной 79 °С. Рибосомы грамцоложительной термофильной бациллы Вас. stearothermophilus денатурировались при температуре 72 °С.
Химический состав рибосом Т. aquaticus следующий: 59% белка и 41% РНК. По процентному содержанию этих компонентов рибосомы Т. aquaticus заметно отличались от рибосом грамотрицательной бактерии В. coli, которая состояла из 41% белка и 59% РНК. Увеличение содержания белка, по-видимому, обеспечивает повышенную термостабильность рибосом культуры Т. aquaticus.
При изучении нуклеотидного состава рибо-сомальной РНК, выделенной из клеток Т. aquaticus, Вас. stearothermophilus и В. coli, было выявлено повышенное содержание гуанина и цитозина в молекуле РНК Т. aquaticus и Вас. stearothermophilus по сравнению с РНК, выделенной из В. coli.
Рибосомальная РНК, выделенная из клеток термофильных бактерий, более устойчива к действию рибосомальной рибонуклеазы, чем РНК, выделенная из мезофильных форм микроорганизмов.
Интересно было сравнить термоустойчивость рибосом Т. aquaticus и В. coli с терморезистентностью выделенных из этих рибосом рибосомальных РНК. Отмечено, что, хотя рибосо-мальная РНК Т. aquaticus более термоустойчива, чем р-РНК В. coli, обе изолированные из рибосом р-РНК были значительно менее термостабильными, чем интактные рибосомы этих же микроорганизмов.
Те клеточные элементы, которые у термофилов термолабильны (транспортная РНК), способны быстро восстанавливаться после их разрушения или инактивации. В. Бабелаи И. Холдстворт установили, что оборачиваемость т-РНК в клетках термофилов гораздо выше, чем у мезофильных форм; этим обеспечивается более высокая скорость синтеза протеинов термофилами. По данным С. М. Фридман и И. Б. Вайнштейн, фермент аминоацил-т-РНК-синтетаза, выделенный из термофильной бактерии Вас. stearothermophilus, обладает высокой термостабильностыо, сама же т-РНК термолабильна.
Таким образом, клетки термофилов способны к быстрому ресинтезу разрушенных и инак-тивированных клеточных компонентов. С другой стороны, т-РНК экстремально-термофильной бактерии Т. aquaticus была более термо-стабилъной и не подвергалась денатурации при 68 °С, в то время как т-РНК В. coli была стабильной только до 55 °С.
Вероятно, высокая термостабильность т-РНК Т. aquaticus, так же как и р-РНК, связана с повышенным содержанием в ее молекуле гуанина и цитозина в сравнении с более низкими концентрациями этих оснований в т-РНК В. coli и Вас. stearothermophilus.
Различные ферментные системы, а также отдельные ферменты термофилов неодинаково устойчивы к действию высокой температуры. Наиболее устойчивы гидролитические ферменты. По степени термостабильности Л. Л. К э м п -белл и Б. Пейс разделяют ферменты на три группы. Среди них наиболее стабильными являются а-амилаза, протеаза, ферменты, активирующие аминокислоты.
Высокой терморезистентностью отличаются аминоацилирующие ферменты, принимающие участие в синтезе белка экстремально-термофильной бактерии Т. aquaticus. Эти ферменты были более термостабильными и биологически активными при оптимальной температуре их действия, равной 70°С, чем те же ферменты В. coli при 45 °С. На высокую степень биологической активности аминоацилсинтетаз Т. aquaticus указывает тот факт, что процесс аминоацилирования у Т. aquaticus заканчивается в первые 3 мин, а у В. coli полное ами-ноацилирование наступало через 6 мин.
Эти данные согласуются с результатами исследователей, изучавших свойства аминоацилт-РНК-синтетаз облигатно-термофильной бактерии Вас. stearothermophilus. Этот фермент был биологически активен при более высокой температуре (55 °С), чем взятый для сравнения фермент из В. coli (37 °С). Однако некоторые экспериментальные данные свидетельствуют о том, что высокий температурный оптимум для ами-ноацилирования у Вас. stearothermophilus не является необходимым. Б. Бабела и И. Холд-сворт сообщили о потере 50% активности аминоацилсинтетазы Вас. stearothermophilus после инкубирования этого фермента при температуре 60 °С в течение 10 мин. Термостабильность аминоацилсинтетазы Вас. stearothermophilus оказалась низкой при температуре, близкой к оптимальной температуре роста указанного микроорганизма. В противоположность этому у Т. aquations обнаружена хорошая корреляция между температурным оптимумом ами-ноацилирования и оптимальной температурой роста, равной 70 °С.
Из экстремально-термофильной бактерии (штамм АТ-62) был выделен фермент аспарто-киназа. Этот фермент катализирует образование таких аминокислот, как лизин, треонин и изолейцин. Аспартокиназа очень термостабильна. Максимально активен этот фермент при 70 °С, что коррелирует с оптимальной температурой роста указанного штамма бактерии.
Фермент фруктозо-1,6-дифосфатальдолаза был выделен из бактерии Т. aquaticus. Температурный оптимум действия этого фермента около 95 °С; он стабилен при 80 °С в течение 1 ч в отсутствие субстрата и при низкой концентрации белка. Значительный интерес представляет сравнение терморезистентности фрук-тозо-1,6-дифосфатальдолаз, выделенных из клеток Т. aquaticus и Вас. stearothermophilus. Оба фермента термостабильны, но фермент, выделенный из Т. aquaticus, был более устойчив к прогреву. Так, указанный фермент, выделенный из Вас. stearothermophilus, инактиви-ровался при 75 °С, а фермент, выделенный из клеток Т. aquaticus УТ-1, был стабилен при 97 °С.
Ферменты гликолитического превращения глюкозы — фосфофруктокиназа и фосфоглюко-мутаза — были выделены японскими исследователями из клеток экстремально-термофильной бактерии Flavobacterium thermophilus НВ-8. Эти ферменты термостабильны. Так, фосфофруктокиназа не инактивировалась в течение 1 ч при 80 °С, и за то же время потеря активности составила всего 10% при температуре прогрева, равной 90 °С. Фосфоглюкомута-за была стабильна при 85 °С. Такая высокая термоустойчивость этих ферментов — одно из характерных свойств экстремально-термофильных бактерий. Эти ферменты играют важную роль в регуляции углеводного обмена при высоких температурах.
Среди различных гидролитических ферментов, выделенных из термофильных микроорганизмов, развивающихся при 55—65 °С, наиболее изучена а-амилаза. Она не инактивиру-ется после 24-часовой выдержки при 65 — 70 °С. Активность этого фермента утрачивалась на 29% от первоначальной лишь после суточной выдержки при 85 °С. Высокая термостабильность а-амилазы обусловлена повышенным содержанием ионов кальция в ее молекуле, а также повышенной стойкостью к агентам, разрушающим водородные связи, а-амилаза была выделена из облигатно-термофильной бактерии Вас. circulans 186 с оптимальной температурой развития 56 °С и термотолерантного штамма Вас. subtilis 110 с оптимальной температурой развития 50 °С. Наиболее высокой термостабильностью обладает а-амилаза облигат-но-термофильного штамма Вас. circulans 186. Инкубация этого фермента при 65 °С в течение 5 ч приводит к снижению активности на 85%. Полная потеря активности в этих же условиях наблюдалась у а-амилазы, выделенной из мезо-фильной культуры Вас. subtilis.
Обнаружены различия в аминокислотном составе а-амилазы термофильных и мезофильных форм микробов. Содержание глютаминовой и аспарагиновой аминокислот в я-амилазах термофильных штаммов больше, чем в а-ами-лазах мезофилов. Как известно, эти аминокислоты могут легко связываться с ионами кальция.
Многие исследователи предполагают, что кальций стабилизирует вторичную и третичную структуру а-амилазы. Было установлено, что существует зависимость между оптимальной температурой роста бактерий и молекулярной массой выделенной из них а-амилазы. Так, а-амилаза, выделенная из Вас. circulans штамм 186, имеет молекулярный вес около 17 000; у а-амилазы, выделенной из Вас. subtilis штамм 110 (с пониженным температурным оптимумом развития), молекулярный вес оказался 28 000—30 000; а-амилаза, выделенная из различных форм мезофильных бактерий Вас. subtilis, имеет молекулярный вес 48 000— 50 000.
В настоящее время интенсивно изучаются особенности термостабильной протеазы. Проводятся глубокие исследования по изучению структуры и физико-химических свойств этого фермента, выделенного из различных форы термофильных микроорганизмов.
Так, был выделен термофильный актиномицет Micromonospora vulgaris штамм 42, развивающийся при 53—55 °С и активно синтезирующий протеазу. Изучены условия культивирования актиномицета и свойства протеолитических ферментов, которые он синтезирует. Выяснилось, что протеаза имеет четыре оптимума, в зависимости от рН раствора. Полученные в чистом виде препараты нейтральных протеаз из культуры актиномицета обладали повышенной термостабильностью и отличались от нейтральных протеаз, выделенных из мезофиль-ных микроорганизмов, рядом физико-химических свойств и специфичностью действия. Из указанного препарата протеаз было выделено два гомогенных препарата, которые заметно отличались своими свойствами. Молекулярный вес одной из протеаз около 50 000, температурный оптимум роста 75 °С. Другая протеаза имела молекулярный вес около 30000, температурный оптимум роста 62—65 °С. Наиболее высокой активностью обладала протеаза с молекулярным весом 50 000.
Многие свойства облигатно-термофильных микроорганизмов закреплены наследственно, и эти формы не могут существовать при перенесении их в обычные температурные условия.
Споры термофильных микроорганизмов обладают значительно большей термоустойчивостью, чем споры мезофильных форм. Предполагают, что повышенная устойчивость спор термофилов к высокой температуре объясняется увеличенным содержанием в них дипико-линовой кислоты, а также уменьшением отношения количества магния к кальцию. Отмечено увеличение содержания в спорах термофильной бактерии Вас. stearothermophilus ли-пидов.
Термофильные бактерии широко распространены в природе. Их можно выделить из почвы, торфа, ила, воды, компоста, навоза и т. д. Но не всегда эти организмы развиваются в термофильных зонах. Однако наиболее интенсивное развитие термофилов наблюдается в местах, подвергающихся воздействию высоких температур. Особенно это касается бактерий, которые не способны развиваться при температуре ниже 40 °С. Эти бактерии называются обли-гатно-термофильными, имеют температурный оптимум роста 55—65 °С и выше.
В значительных количествах термофильные бактерии обнаруживаются в огородных и полевых почвах, куда они попадают в основном вместе с органическими удобрениями. Было показано, что в окультуренных почвах термофилов довольно много, а в необработанных почвах их почти нет. Эти наблюдения дали возможность использовать термофильные микроорганизмы в качестве показателей степени окультуренности почв. Однако термофильные микроорганизмы были обнаружены даже в почвах и водах Крайнего Севера.
По сравнению с почвой такие субстраты, как сточные воды, компост, навоз, самонагревающиеся торф и сено, содержат большее количество термофильных бактерий. Самонагревание сена и торфа, которое иногда приводит к пожару, в значительной мере обусловливается развитием термофилов. Наличие термофильных микроорганизмов в кишечном тракте и экскрементах животных и человека отмечено многими исследователями.
Интересно отметить, что такие продукты, как нефть и входящие в ее состав вещества, также могут усваиваться термофильными микроорганизмами.
Широко распространены термофильные суль-фатвосстанавливающие бактерии, оптимальная температура роста которых достигает 55—60 °С. Известны также термофильные бактерии, окисляющие различные соединения серы до молекулярной серы и серной кислоты. Горячие источники, богатые сероводородом, изобилуют термофильными видами тионовых бактерий. Из Брагунских терм (Северный Кавказ) с температурой 89—90 °С выделена тиобактерия Thiobacillus thermophilica Imschenetskii. Оптимальная температура роста этой бактерии 55 — 60 °С, максимальная — около 80 °С, минимальная 40 °С.
С. И. Кузнецов в 1955 г. обнаружил, что в источниках Камчатки с температурой 90—98 °С развиваются термофильные бактерии. В 1967 —1971 гг. американскими исследователями были выделены из субтермальных вод с температурой 85—89 °С бактерии, не образующие спор, которые не способны развиваться при температуре ниже 40—45 °С, они относятся к виду Thermus aquaticus. Температурный оптимум развития этих бактерий 70—80 °С.
Термофильные анаэробные бактерии рода Clostridium были обнаружены во всех обследованных пробах воды и почв, взятых в термальных зонах, а также в компостах.
Экстремально-термофильные бактерии
В последние годы американскими исследователями Т. Д. Броком и X. Фризом была описана новая группа бактерий — экстремальные термофилы, в которую вошли виды, способные развиваться при крайне высоких (экстремальных) температурах. Эти формы выделены из различных термальных источников, вода которых имела температуру 85—95 °С и слабощелочную реакцию.
Бактерии — неподвижные грамотрицательные палочки; оптимальная температура развития 70 °С. Они описаны как виды нового рода термус (Thermus). Позднее подобные формы бактерий были найдены многими исследователями.
Как известно, при повышении температуры изменяется характер воздействия на клетку ряда физических и химических факторов. Так, при высокой температуре уменьшаются вязкость и увеличивается степень ионизации воды, уменьшается растворимость кислорода и других газов в водной среде, ускоряется течение химических реакций и т. д. Брок указывает, что в подобных условиях существования происходит эволюционная адаптация, при которой организм полностью зависит от определенного значения решающего фактора, летального (губительно действующего) на другие микроорганизмы.
По мере повышения температуры разнообразие обнаруживаемых групп различных микроорганизмов сужается. Первыми исчезают наиболее сложные по строению организмы. Высокие температуры хорошо переносят только бактерии.
В большинстве случаев при экстремальных условиях (в частности, при наиболее высоких температурах, 80—90 °С) существующие микроорганизмы приближаются к чистой культуре в местах обитания, т. е. к одному виду. Необходимо отметить, что в источниках с высокой температурой часто обнаруживаются наряду с указанными бактериями также и термофильные водоросли. Такие водоросли благодаря их способности фиксировать углекислый газ и молекулярный азот могут развиваться в воде горячих источников с низким содержанием органических веществ. По-видимому, углерод и азот органических соединений, содержащихся в водорослях, ассимилируется бактериями. Эта зависимость развития бактерий от контакта с водорослями может быть, по мнению Б рока, настолько велика, что без водорослей в горячих источниках с низким содержанием органических веществ рост бактерий становился бы невозможным.
Способность неспорообразующих бактерий, обитающих в горячих источниках, существовать в природе при температуре от 40 до 93 °С и выше дает основание для выделения этих микроорганизмов в новую группу экстремально-термофильных бактерий. Сверхтермофиль-ность этих бактерий по сравнению с облигатно-термофильными бациллами характеризуется более высокими температурными параметрами роста. Указанная группа экстремально-термофильных бактерий специфическая, что обусловлено местом обитания этих микроорганизмов.
Thermus aquaticus широко распространен в природе. Бактерии этого нового вида плохо развиваются при 55 °С, температурный минимум роста 42 °С, при 40 °С рост не отмечен. Оптимальная температура роста 70 °С, максимальная 79 °С. Эти бактерии являются обли-гатными аэробами. Оптимум рН роста 7,5—7,8. Культура имеет желтую или оранжевую окраску. Размер палочек 0,5 X 5 — 10 мкм.
При температурах выше и ниже оптимальной рост происходит при образовании длинных нитей.
Из различных горячих источников были выделены бактерии подобного вида, способные расти при 84 °С и даже при 91 °С; клетки их бледно-желтого цвета. Выделены также бактерии, не образующие пигмента; температурный диапазон развития их от 40 до 80 °С, оптимальная температура роста 69—71 °С.
Спорообразующие аэробные термофильные бактерии
Эти формы бактерий, как указывалось выше, широко распространены в природе. Многие мезофильные виды бацилл имеют своих аналогов среди термофилов.
Рис. 189. Термофильная бактерия Вас. stearotliermophilus. Увел. X 3000.
В настоящее время известно всего лишь два самостоятельных вида термофильных спо-рообразующих бактерий — Вас. stearother-mophilus и Вас. coagulans. Клетки Вас. stearo-thermophilus представлены на рисунке 189.
Процессы роста и размножения у термофильных микроорганизмов проходят с большей скоростью, чем у мезофильных форм микробов. У облигатно-термофильных бактерий, которые не способны развиваться при температурах ниже 37—40 °С и имеют оптимальную температуру роста 55—65 °С, весь цикл развития проходит за 5—8 ч.
Скорость роста мезофильных культур по сравнению со скоростью роста термофилов значительно ниже.
Продолжительность фазы интенсивного роста (лаг-фаза) облигатно-термофильных бактерий чрезвычайно коротка. Продолжительность лаг-фазы термофильных микроорганизмов меньше, чем мезофильных. Так, у Вас. coagulans наименьшая продолжительность лаг-фазы равна 15 мин при 65 °С роста культуры и 20— 25 мин при 55 °С.
Максимальное количество клеток, накапливаемое в культурах облигатно-термофиль-ных бактерий, колеблется в пределах 109— 1,3-109 на 1 мл при 55—60 °С и снижается до 6.Ю8-8-108 при 65-70 °С.
Влияние аэрации и перемешивания на рост и развитие аэробных термофильных бактерий
Одним из важнейших факторов, определяющих рост и развитие термофильных микроорганизмов, является скорость поступления кислорода и его концентрация в культуральной среде. Степень ограничения роста аэробных организмов при недостатке кислорода зависит от температуры выращивания. Растворимость кислорода в воде увеличивается с понижением температуры, поэтому рост микроорганизмов при более низких температурах не ограничивается содержанием кислорода в такой степени, как в случае инкубации при высоких температурах. Этим и объясняется тот факт, что общий урожай организмов, выращенных при низких температурах, часто оказывается выше, чем урожай микроорганизмов, выращенных при более высоких температурах, хотя скорость роста в последнем случае может быть больше.
Процессы метаболизма в клетках термофилов протекают с гораздо большей скоростью, чем в клетках мезОфилов. Поэтому концентрация растворенного в среде кислорода может явиться фактором, лимитирующим рост термофильных микроорганизмов. Однако при культивировании термофильных микроорганизмов на богатых естественных средах в условиях интенсивной аэрации организмы могут и не испытывать недостатка в растворенном кислороде. Но при выращивании термофилов на синтетических средах количество растворенного кислорода начинает выступать в роли решающего фактора.
Очень интересны опыты Р. Дж. Доунея. Этот исследователь показал, что при повышенной температуре необходимо насыщение среды кислородом под давлением, равным 1 атм. В этих условиях растворимость кислорода увеличивается. Так, при 60 °С концентрация кислорода была равна 139 микромолям, это значение близко к концентрации кислорода в среде для развития мезофильных форм при обычных температурах (143 — 240 микромолей). Выращивая Вас. stearothermophilus на полноценной питательной среде с повышенным содержанием кислорода при 60 °С, Доунею удалось получить большую биомассу этих бактерий. Таким образом, выяснилось, что биомасса клеток является функцией концентрации кислорода в среде и максимальное ее количество достигается при растворении кислорода в среде под давлением, равным 1 атм. Дальнейшее увеличение концентрации кислорода замедляет рост бактерий.
Процесс дыхания у термофильных микроорганизмов осуществляется гораздо интенсивней, чем у мезофилов. В лаборатории Л. Г. Логиновой был отмечен рштересный факт, ранее не описанный в литературе. При ускорении процесса дыхания с повышением температуры культивирования в клетках термофильных микроорганизмов заметно увеличивалось количество цитохромов. Особенно значительно оно возрастало в клетках облигатно-термофильных бактерий Вас. stearothermophilus, Вас. circu-lans, Вас. megaterium, Вас. brevis при температуре выращивания 65 °С. При этой температуре количество цитохромов возросло примерно в 2—2,5 раза по сравнению с их количеством в клетках бактерий, выращенных при температуре 55 °С.
Интересно отметить, что некоторое увеличение количества цитохромов при повышении температуры роста также наблюдалось и у мезофилов, но оно было значительно слабее, чем у термофильных форм микробов.
Потребность термофилов в питательных веществах зависит от температуры их роста. Л. Л. Кэмпбелл и Б. Пейс разделили по этому признаку термофильные спорообразующие бактерии на три группы. К первой группе относятся термофильные бактерии, потребность в питательных веществах которых не зависит от температуры. Бактерии второй группы нуждаются в дополнительном питании при повышении температуры выращивания, .а третьей группы — при понижении температуры.
Анаэробные термофильные бактерии
Значительную часть спороносных термофильных бактерий составляют анаэробные виды. Известны облигатно-термофильные масляно-кислые, целлюлозные, десульфурирующие и метанобразующие бактерии.
Термофильные целлюлозные бактерии. Этих бактерий обнаруживают обычно в компостах, разлагающихся растительных отбросах, илах и т. д. В таких субстратах эти бактерии наряду с термофильными целлюлозоразлагающими грибами и актиномицетами находят благоприятные условия для своего развития. Когда температура поднимается до 60—65 °С, физиологическая активность грибов и актиномицетов резко снижается и разрушение клетчатки полностью осуществляется бактериями.
Развитие целлюлозных бактерий можно заметить сначала по газообразованию, затем по разрушению клетчатки (фильтровальной бумаги) и появлению желто-оранжевого пигмента.
Чистые культуры этих бактерий получить довольно трудно, и это удавалось немногим исследователям. Типичным мезофильным представителем является бактеррш Bacillus omeli-anskii, названная в честь В. Л. Омелянского, впервые описавшего этот микроорганизм. В качестве типичного термофильного представителя можно назвать Clostridium thermocellulaseum. Описания мезофильных и термофильных видов указанных бактерий тождественны, поэтому А. А. Имшенецкий считает, что они представляют один вид. При этом термофилы могут рассматриваться как варианты мезофилов.
Морфологически целлюлозные бактерии представляют собой тонкие, прямые или слегка изогнутые палочки, часто с округлыми спорами на концах клеток. Палочки подвижны, жгутики расположены по всей поверхности клетки.
Продуктами обмена веществ анаэробных целлюлозных бактерий, выделяющимися в окружающую среду, являются водород, углекислый газ, этиловый спирт, глицерин, муравьиная, уксусная, молочная, янтарная, яблочная и фумаровая кислоты. Присутствуют также следы ацетальдегида, иногда — масляная кислота. Эти бактерии разлагают целлюлозу до низкомолекулярных углеводов, главным образом до целлобиозы и глюкозы. Гидролиз целлюлозы сопровождается появлением в среде фермента целлюлазы.
Термофильные метановые бактерии могут сопутствовать анаэробным целлюлозным бактериям или культивироваться совместно с ними. В начале этого века Ц. К о-олхаасом впервые были описаны термофильные метановые бактерии с температурными границами развития 45—69 °С. Наиболее типичными видами являются Methanobacterium soehngenii и Methanobacillus omelianskii. Эти бактерии представляют собой тонкие, прямые или слегка изогнутые неспороносные палочки.
Термофильные десульфурирующие бактерии нередко сопутствуют термофильным целлюлозным бактериям. При высоких температурах восстановление сульфатов осуществляется бактерией Desulfotomaculum nigrificans, которая представляет собой палочки с округлыми концами, иногда чечевицевидные и вздутые, подвижные, пери-трихи. Споры овальные, субтерминальные или терминальные.
Термофильные лучистые грибки (актиномицеты)
К термофильным лучистым грибкам относятся актиномицеты различных систематических групп, обладающие способностью развиваться при высоких температурах (50—60 °С), независимо от температурного минимума их роста.
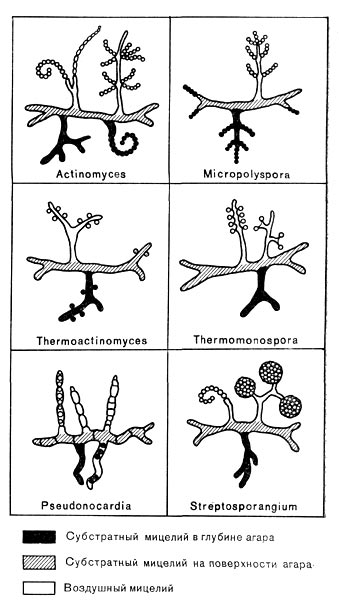
Рис. 190. Схематическое изображение вегетативного и спороносного мицелия различных родов термофильных актиномпцетов.
Среди них встречаются актиномицеты, способные расти при 60—70 °С. Лучистые грибки, развивающиеся при обычных температурах (25 — 30 °С), не растут при температуре 50 °С и выше.
Количественный учет термофильных актино-мицетов в почвах и компостах был проведен В. Ваксманом с сотрудниками в 1939 г. Термофильные актиномицеты были обнаружены во всех почвах и во все сезоны года. Особенно много их в почвах, удобренных навозом (в среднем 200 000 на 1 г в весенних и летних пробах). Зимой термофильные актиномицеты составляли 10—15% от всей термофильной микрофлоры; весной и летом 70—90%. Количество термофильных лучистых грибков не зависит от географической закономерности, а определяется экологическими факторами, в частности типом почвы и степенью ее окультуреннооти.
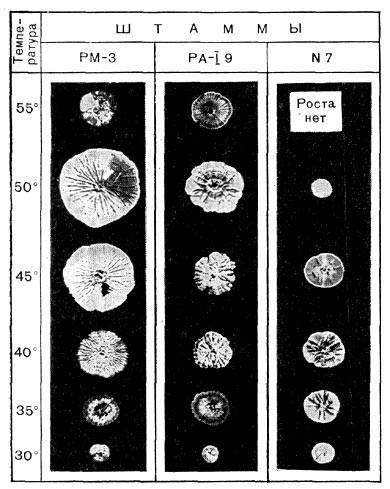
Рис. 191. Гигантские колонии различных актиномицетов при разных температурах. Уменьшено в 2,5 раза.
Н. Д. Красильников (1970) указывает, что в пределах одного семейства и рода (например, Micromonospora) могут быть как термофильные, так и мезофильные культуры; несмотря на довольно стойкий признак термофильности, видимо, нецелесообразно ориентироваться на него при характеристике родов или более крупных таксономических единиц.
Актиномицеты, способные развиваться при 40—60 СС, встречаются среди представителей различных родов, но чаще всего среди культур рода Micromonospora. Ранее признак термофильности отражали в некоторых родовых названиях актиномицетов, растущих при повышенных температурах (рис. 190). При идентификации термофильных актиномицетов используют, как обычно, культурал.ьные признаки.
Большинство известных термофильных лучистых грибков быстро гидролизуют крахмал, свертывают и пептонизируют молоко, разжижают желатин и т. д., что свидетельствует о высокой ферментативной активности и может быть использовано в практике. Однако эти культуральные свойства лабильны и поэтому, с точки зрения многих исследователей, не могут быть основными критериями при определении вида.
Другие культуральные свойства, такие, как восстановление нитратов, образование сероводорода, в большей степени отражают физиологические особенности микроорганизма, более стабильны и, следовательно, играют существенную роль при идентификации вообще и термофильных актиномицетов в частности.
Для выделения термофильных актиномицетов разными авторами использовались разные методики. Выделение этих микроорганизмов хорошо удается на крахмальном агаре, крах-мально-аммиачно-сульфатпом агаре, а также мясо-пептонном агаре (МПА). Лучшим для этой цели оказался крахмально-аммиачно-сульфатный агар, на котором наблюдается слабый рост более требовательных к среде термофильных бактерий и, наоборот, хороший рост термофильных актиномицетов. Наиболее подходящая температура для выделения 55—60 °С.
Культивируют термофильные актиномицеты на самых различных средах.
Для получения хорошего роста и споруляции этих микроорганизмов обычно используют крахмал и неочищенную мальтозу. К. Е. Эриксон отмечает, что Micromonospora vulgaris хорошо растет при использовании следующих источников азота: пептона, трип-тона, гидролизата, казеина. Многие термофильные актиномицеты нуждаются в дополнительных компонентах среды, представляющих собой смеси аминокислот, витаминов, пуринов и пиримидинов.
Термофильные актиномицеты обладают большой скоростью роста. Их жизненный цикл проходит гораздо быстрее, чем у мезофильных штаммов. Термофильные актиномицеты образуют разветвленный мицелий из гиф, диаметр которых от 0,2 до 1 мкм. На твердых средах они растут в виде плоских колоний, достигающих 3—4 см в диаметре, а иногда и до 6—8 см (рис. 191), часто покрытых налетом, состоящим из воздушных гиф со спорами. Термофильные актиномицеты образуют воздушный и субстратный мицелии. Гифы воздушного мицелия без спор термофильных лучистых грибков, как правило, белоснежно-белого цвета. Воздушный мицелий со спорами или сохраняет белый цвет, или приобретает темно-серый оттенок. Серо-зеленые, голубые и желтые штаммы встречаются реже.
У некоторых термофильных актиномицетов в процессе развития изменяется цвет колоний на агаре от белоснежно-белого до желтого, грязно-зеленоватого, коричневого, красноватого и даже черного. Многие представители термофильных лучистых грибков образуют растворимый пигмент, который проникает в среду и окрашивает ее в яркие цвета.
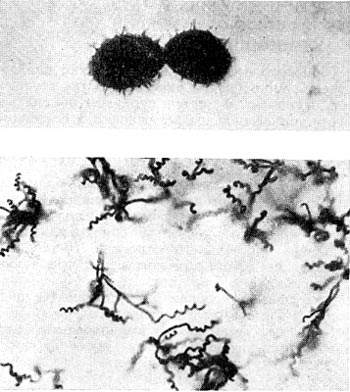
Рис. 192. Термофильный актиномпцет Actinomyces diastaticus, штамм 7. Вверху — споры с шиповидной поверхностью (увел. X 10 000); внизу - форма спороносцев (увел. X 900).
Разные исследователи описывают наличие у термофильных актиномицетов прямых или спирально закрученных спороносцев (рис. 192). Имеются также формы с одиночными, парными спорами и спорами в виде цепочек, образованных на коротких веточках вегетативного воздушного мицелия. Споры всех термофильных актиномицетов круглые или эллипсоидальные (рис. 192) диаметром от 0,6 до 1,4 мкм. В них обнаружена дипиколиновая кислота, а также кальций и в меньшей степени магний, которые, как известно, обусловливают высокую термоустойчивость спор бактерий.
Изучение тонкого строения спор ряда актиномицетов позволило исследователям сделать вывод, что некоторые термофильные актиномицеты образуют споры, близкие по строению к спорам бактерий родов Bacillus и Clostridium.
Обмен веществ у термофильных микроорганизмов происходит более интенсивно, чем у мезофильных. Об этом свидетельствуют экспериментальные данные о более высокой ферментативной активности термофилов и о повышенном количественном содержании в клетках термофильных микроорганизмов некоторых ферментов.
Так, при изучении цитохромов в клетках мезофильных и термофильных представителей различных родов актиномицетов было обнаружено, что у некоторых термофильных актиномицетов (Thermoactinomyces sp., Actinobi-fida dichotomica, Mycropolyspora sp.) оказалось больше цитохромов типов с и а; очевидно, некоторые участки цепи переноса электронов (в частности, цитохромная система) термофильных штаммов могут значительно отличаться от таковых у мезофильных форм.
Нередки случаи, когда термофильные актиномицеты способны образовывать в больших количествах экзоферменты, действующие на различные субстраты, что может быть использовано в практике. Так, выделен термофильный актиномицет Act. diastaticus штамм 7 (рис. 192), образующий целлюлозолитические и гемицеллюлозолитические ферменты. Однако образование подобных экзоферментов не является свойством, специфическим для термофилов.
С целью выяснения биохимических особенностей термофильных актиномицетов многие исследователи изучали состав нуклеотидов ДНК как термофильных, так и мезофильных видов. Результаты исследований дают возможность полагать, что термофильные актиномицеты содержат сравнительно меньше ГЦ (гуанин + цитозин) в составе ДНК, чем мезофиль-ные штаммы. Этот вопрос требует дальнейших исследований. В биомассе актиномицетов, выросших при высокой температуре (55—57 °С), содержится в 2 раза больше свободных нуклеотидов; количество нуклеиновых кислот, наоборот, падает. Свободные нуклеотиды, видимо, играют более значительную роль в интенсификации обмена веществ у термофилов по сравнению с мезофильными микроорганизмами.
Есть данные о том, что термофильные штаммы рода Actinomyces не чувствительны к фагам, активным против мезофильных штаммов того же рода, хотя существенных биохимических различий в составе клеточных стенок у мезофильных и термофильных актиномицетов не обнаружено.
Практическое значение термофильных бактений и актиномицетов
Термофильные бактерии используют для получения микробной биомассы, очистки сточных вод. Ценными являются продукты обмена веществ термофилов, выделяющиеся в окружающую среду. Эти микроорганизмы продуцируют такие физиологически активные вещества, как антибиотики, витамины, ферменты.
Обычно для получения микробной биомассы используют термотолерантные дрожжи. Их выращивают на средах, содержащих углеводы (сусловые среды), некоторые спирты или углеводороды нормального строения (н-алканы). В последнее время для этих целей применяют и термофильные бактерии.
Выросшая микробная биомасса вполне полноценна в пищевом отношении: содержит 40— 60% белка, незаменимые аминокислоты, разнообразные витамины. Высушенная биомасса (в виде муки) — белково-витаминный концентрат (БВК) — в небольшом количестве добавляется к пищевому рациону животных.
Продукты обмена веществ термофильных бактерий нашли широкое применение в промышленности. Так, молочнокислые бактерии Bact. delbruckii используются как активные кислото-образователи. Еще в 1923 г. В. М. Шапошникову и А. Я. Мантейфель удалось наладить производство молочной кислоты с помощью термофильных бактерий. Ряд термофильных молочнокислых бактерий применяется в молочной промышленности для получения высококачественного творога.
Из различных физиологически активных веществ, продуцируемых термофильными микроорганизмами и используемых в практике, огромное значение имеют ферменты.
Список ферментов, продуцируемых термофильными микроорганизмами и применяемых в промышленности, очень обширен.
Так, в текстильной промышленности А. А. Имшенецким с сотрудниками была применена амилаза. Этот фермент образовывала термофильная бактерия Вас. diastaticus. Амилаза используется и в спиртовой промышленности для размягчения зерновых и картофельных сред при высоких температурах.
Протеазы применяют для переработки сырья животного происхождения (обезволашивания кожи, получения клея и т. д.). Выделены и изучены термофильные актиномицеты, активно образующие протеолитические и амилолитические ферменты, комплекс целлюлозе- и гемицеллюлозолитических ферментов.
Для получения целлюлолитических ферментов пытались использовать термофильные анаэробные бактерии и актиномицеты. С помощью целлюлолитических ферментов можно повысить питательную ценность грубых кормов для животных, осахаривать сульфатную целлюлозу до глюкозы, получая таким образом из непищевого сырья ценный питательный продукт. Эти ферменты способны расщеплять полисахариды одревесневших, растительных материалов (шелуха злаковых культур, подсолнечная лузга) до Сахаров (глюкозы и ксилозы). Следовательно, открывается возможность замены кислотного гидролиза древесины и различных отходов ферментативным гидролизом. При этом полностью ликвидируются расход минеральных кислот и необходимость применения высоких температур и давлений при превращении целлюлозы в сахара.
Чешские ученые используют термофильные грибы и актиномицеты, которые продуцируют разлагающие целлюлозу ферменты, для воздействия на различные субстраты.
Л. Г. Логиновой выделен термофильный актиномицет Micromonospora vulgaris штамм РА-П4, образующий комплекс литических ферментов. Эти ферменты разрушают клеточные стенки различных бактерий и дрожжей (мертвых и живых). Литические ферменты могут найти применение в различных областях народного хозяйства: в повышении усвояемости и питательной ценности кормовой микробной биомассы, в борьбе с бактериальными инфекциями человека и животных и в других областях.
Термофильные бактерии издавна применяются для очистки сточных вод. Интерес к метановому брожению резко возрос, когда была обнаружена способность бактерий продуцировать витамин В12. В. Н. Букин показал возможность получения этого ценного витамина при сбраживании термофильными метановыми бактериями ацетоно-бутиловой барды. Одновременно может быть собран выделяющийся при этом метан (10—20 м3 на 1 м3 сброженной жидкости).
Термофильные микроорганизмы играют существенную роль в круговороте веществ в природе: в разрушении нефтей и озокеритов, превращениях серы и других процессах.
В ряде промышленных производств термофильные бактерии могут приносить и существенный вред: вызывают заражение сусла на пивоваренных заводах, порчу консервов (особенно анаэробные бактерии), сгущенного стерилизованного молока, сахарных сиропов. Поэтому необходимы надежные методы стерилизации.
Как для культивирования полезных форм термофильных бактерий, так и для методов борьбы с вредными для определенных процессов видами необходимо вести глубокое изучение морфологических, физиологических и биохимических особенностей этих микроорганизмов.
|
ПОИСК:
|
При использовании материалов сайта активная ссылка обязательна:
http://plant.geoman.ru/ 'Растения: виды, места произрастания, полезные свойства'